
Latest recommendations

| Id | Title * | Authors * | Abstract * | Picture * ▲ | Thematic fields * | Recommender | Reviewers | Submission date | |
|---|---|---|---|---|---|---|---|---|---|
01 Jul 2020

Sub-lethal insecticide exposure affects host biting efficiency of Kdr-resistant Anopheles gambiaekdr homozygous resistant An. gambiae displayed enhanced feeding success when exposed to permethrin Insect-Treated NetsRecommended by Adrian Diaz based on reviews by Thomas Guillemaud, Niels Verhulst, Etienne Bilgo and 1 anonymous reviewerMalaria is a vector-borne parasitic disease found in 91 countries with an estimated of 228 million cases occurred worldwide during 2018. The 93% (213 million) of those cases were reported in the African Region (WHO 2019). Six species of Plasmodium parasites can produce the disease but only P. falciparum and P. vivax are the predominant species globally. More than 40 species of Anopheles mosquitoes are important malaria vectors (Asley et al. 2018). Intrinsic (genetic background, parasite susceptibility) and extrinsic (feeding host preference, host diversity and availability, mosquito abundance) factors affect the capacity of mosquitoes to vector the disease (Macdonald 1952). Malaria is prevented by chemoprophylaxis, vaccination, bite-avoidance and vector-control measures. The mainstays of vector control are long-lasting insecticide (pyrethroid) treated nets and indoor residual spraying with insecticides (Asley et al. 2018). The widespread use of pyrethroid insecticides forced the emergence of insecticide resistance in malaria vectors reducing the insecticidal effect. Mosquitoes can modify their behaviour avoiding insecticide contact and so potentially reducing vector control tools efficacy. In this sense, Diop et al. (2020) investigated whether pre-exposure to an Insecticide-Treated Net (ITN) modulates the mosquito ability to take a blood meal in Anopheles gambiae. By means of video recording experiments the authors analyzed how the feeding/bitting behaviour was affected by kdr mutation genotypes (homozygous susceptible – SS-, heterozygotes -RS- and homozygous resistant -RR-) when exposed to two different insecticides (permethrin and deltamethrin). According to the results, the blood-feeding success did not differ between the three genotypes in the absence of insecticide exposure. However, authors observed differences in the feeding duration and blood meal size. In example, RR mosquitoes spent less time taking their blood meal than RS and SS. On the other hand, RS mosquitoes took higher blood volumes than RR females. These differences can affect the mosquito fitness by decreasing/increasing the likelihood to be killed by the host defensive behavior or increase the oogenesis so enhancing fecundity. Regarding the effect of exposition to insecticides authors detected a strong relationship between kdr genotype and Knock Down (KD) phenotype when mosquitoes were exposed to Permethrin. Previously, the authors have evidenced that RR mosquitoes prefer a host protected by a permethrin-treated net rather than an untreated net and that heterozygotes RS mosquitoes have a remarkable ability to find a hole into a bet net (Diop et al. 2015, Porciani et al. 2017). With data here obtained, they demonstrated that kdr homozygous resistant An. gambiae displayed enhanced feeding success when exposed to permethrin ITN. The changes observed in the feeding/biting mosquito behaviour can affect their fitness shaping the evolution of the insecticide resistance in mosquitoes’ natural populations. Moreover, this may also alter parasite transmission dynamics by modifying vector/host interactions and so vector capacity. References World Health Organization (2019). World malaria report 2019. Geneva: World Health Organization; 2019. ISBN 978-92-4-156572-1 | Sub-lethal insecticide exposure affects host biting efficiency of Kdr-resistant Anopheles gambiae | Malal Mamadou Diop, Fabrice Chandre, Marie Rossignol, Angelique Porciani, Mathieu Chateau, Nicolas Moiroux, Cedric Pennetier | <p>The massive use of insecticide-treated nets (ITNs) has drastically changed the environment for malaria vector mosquitoes, challenging their host-seeking behaviour and biting success. Here, we investigated the effect of a brief exposure to an IT... | | Behavior, Ecology, Evolution, Medical entomology, Pesticide resistance | Adrian Diaz | Anonymous | 2019-05-29 19:40:25 | |
25 Aug 2022

Improving species conservation plans under IUCN's One Plan Approach using quantitative genetic methodsQuantitative genetics for a more qualitative conservationRecommended by Peter Galbusera based on reviews by Timothée Bonnet and 1 anonymous reviewerGenetic (bio)diversity is one of three recognised levels of biodiversity, besides species and ecosystem diversity. Its importance for species survival and adaptation is increasingly highlighted and its monitoring recommended (e.g. O’Brien et al 2022). Especially the management of ex-situ populations has a long history of taking into account genetic aspects (through pedigree analysis but increasingly also by applying molecular tools). As in-situ and ex-situ efforts are nowadays often aligned (in a One-Plan-Approach), genetic management is becoming more the standard (supported by quickly developing genomic techniques). However, rarely quantitative genetic aspects are raised in this issue, while its relevance cannot be underestimated. Hence, the current manuscript by Sauve et al (2022) is a welcome contribution, in order to improve conservation efforts. The authors give a clear overview on how quantitative genetic analysis can aid the measurement, monitoring, prediction and management of adaptive genetic variation. The main tools are pedigrees (mainly of ex-situ populations) and the Animal Model. The main goal is to prevent adaption to captivity and altered genetics in general (in reintroduction projects). The confounding factors to take into account (like inbreeding, population structure, differences between facilities, sample size and parental/social effects) are well described by the authors. As such, I fully recommend this manuscript for publication, hoping increased interest in quantitative analysis will benefit the quality of species conservation management. References O'Brien D, Laikre L, Hoban S, Bruford MW et al. (2022) Bringing together approaches to reporting on within species genetic diversity. Journal of Applied Ecology, 00, 1–7. https://doi/10.1111/1365-2664.14225 Sauve D., Spero J., Steiner J., Wheeler H., Lynch C., Chabot A.A. (2022) Improving species conservation plans under IUCN’s One Plan Approach using quantitative genetic methods. EcoEvoRxiv, ver. 9 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.32942/osf.io/n3zxp | Improving species conservation plans under IUCN's One Plan Approach using quantitative genetic methods | Drew Sauve, Jane Hudecki, Jessica Steiner, Hazel Wheeler, Colleen Lynch, Amy A. Chabot | <p>Human activities are resulting in altered environmental conditions that are impacting the demography and evolution of species globally. If we wish to prevent anthropogenic extinction and extirpation, we need to improve our ability to restore wi... | | Conservation biology, Ecology, Evolution, Genetics/Genomics | Peter Galbusera | 2022-02-21 10:45:22 | ||
28 Apr 2021
Inference of the worldwide invasion routes of the pinewood nematode Bursaphelenchus xylophilus using approximate Bayesian computation analysisExtracting the maximum historical information on pine wood nematode worldwide invasion from genetic dataRecommended by Stéphane Dupas based on reviews by Aude Gilabert and 1 anonymous reviewer based on reviews by Aude Gilabert and 1 anonymous reviewer
Redistribution of domesticated and non domesticated species by humans profoundly affected earth biogeography and in return human activities. This process accelerated exponentially since human expansion out of Africa, leading to the modern global, highly connected and homogenized, agriculture and trade system (Mack et al. 2000, Jaksic and Castro 2021), that threatens biological diversity and genetic resources. To accompany quarantine and control effort, the reconstruction of invasion routes provides valuable information that help identifying critical nodes and edges in the global networks (Estoup and Guillemaud 2010, Cristescu 2015). Historical records and genetic markers are the two major sources of information of this corpus of knowledge on Anthropocene historical phylogeography. With the advances of molecular genetics tools, the genealogy of these introductions events could be revisited and empowered. Due to their idiosyncrasy and intimate association with the contingency of human trades and activities, understanding the invasion and domestication routes require particular statistical tools (Fraimout et al. 2017). Because it encompasses all these theoretical, ecological and economical implications, I am pleased to recommend the readers of PCI Zoology this article by Mallez et al. (2021) on pine wood nematode invasion route inference from genetic markers using Approximate Bayesian Computation (ABC) methods. Economically and ecologically, this pest, is responsible for killing millions of pines worldwide each year. The results show these damages and the global genetic patterns are due to few events of successful introductions. The authors consider that this low probability of introductions success reinforces the idea that quarantine measures are efficient. This is illustrated in Europe where the pine-worm has been quarantined successfully in the Iberian Peninsula since 1999. Another relevant conclusion is that hybridization between invasive populations have not been observed and implied in the invasion process. Finally the present study reinforced the role of Asiatic bridgehead populations in invasion process including in Europe. Methodologically, for the first time, ABC was applied to this species. A total of 310 individual sequences were added to the Mallez et al. (2015) microsatellite dataset. Fraimoult et al. (2017) showed the interest to apply random forest to improve scenario selection in ABC framework. This method, implemented in the DiYABC software (Collin et al. 2020) for invasion route scenario selection allows to handle more complex scenario alternatives and was used in this study. In this article by Mallez et al. (2021), you will also find a clear illustration of the step-by-step approach to select scenario using ABC techniques (Lombaert et al. 2014). The rationale is to reduce number of scenario to be tested by assuming that most recent invasions cannot be the source of the most ancient invasions and to use posterior results on most ancient routes as prior hypothesis to distinguish following invasions. The other simplification is to perform classical population genetic analysis to characterize genetic units and representative populations prior to invasion routes scenarios selection by ABC. Yet, even when using the most advanced Bayesian inference methods, it is recognized by the authors that the method can be pushed to its statistical power limits. The method is appropriate when population show strong inter-population genetic structure. But the high number of differentiated populations in native area can be problematic since it is generally associated to incomplete sampling scheme. The hypothesis of ghost populations source allowed to bypass this difficulty, but the authors consider simulation studies are needed to assess the joint effect of genetic diversity and number of genetic markers on the inference results in such situation. Also the need to use a stepwise approach to reduce the number of scenario to test has to be considered with caution. Scenarios that are not selected but have non negligible posterior, cannot be ruled out in the constitution of next step scenarios hypotheses. Due to its interest to understand this major facet of Anthropocene, reconstruction of invasion routes should be more considered as a guide to damper biological homogenization process. References Collin, F.-D., Durif, G., Raynal, L., Lombaert, E., Gautier, M., Vitalis, R., Marin, J.-M. and Estoup, A. (2020) Extending Approximate Bayesian Computation with Supervised Machine Learning to infer demographic history from genetic polymorphisms using DIYABC Random Forest. Authorea. doi: https://doi.org/10.22541/au.159480722.26357192 Cristescu, M.E. (2015) Genetic reconstructions of invasion history. Molecular Ecology, 24, 2212–2225. doi: https://doi.org/10.1111/mec.13117 Estoup, A. and Guillemaud, T., (2010) Reconstructing routes of invasion using genetic data: Why, how and so what? Molecular Ecology, 9, 4113-4130. doi: https://doi.org/10.1111/j.1365-294X.2010.04773.x Fraimout, A., Debat, V., Fellous, S., Hufbauer, R.A., Foucaud, J., Pudlo, P., Marin, J.M., Price, D.K., Cattel, J., Chen, X., Deprá, M., Duyck, P.F., Guedot, C., Kenis, M., Kimura, M.T., Loeb, G., Loiseau, A., Martinez-Sañudo, I., Pascual, M., Richmond, M.P., Shearer, P., Singh, N., Tamura, K., Xuéreb, A., Zhang, J., Estoup, A. and Nielsen, R. (2017) Deciphering the routes of invasion of Drosophila suzukii by Means of ABC Random Forest. Molecular Biology and Evolution, 34, 980-996. doi: https://doi.org/10.1093/molbev/msx050 Jaksic, F.M. and Castro, S.A. (2021). Biological Invasions in the Anthropocene, in: Jaksic, F.M., Castro, S.A. (Eds.), Biological Invasions in the South American Anthropocene: Global Causes and Local Impacts. Springer International Publishing, Cham, pp. 19-47. doi: https://doi.org/10.1007/978-3-030-56379-0_2 Lombaert, E., Guillemaud, T., Lundgren, J., Koch, R., Facon, B., Grez, A., Loomans, A., Malausa, T., Nedved, O., Rhule, E., Staverlokk, A., Steenberg, T. and Estoup, A. (2014) Complementarity of statistical treatments to reconstruct worldwide routes of invasion: The case of the Asian ladybird Harmonia axyridis. Molecular Ecology, 23, 5979-5997. doi: https://doi.org/10.1111/mec.12989 Mack, R.N., Simberloff, D., Lonsdale, M.W., Evans, H., Clout, M., Bazzaz, F.A. (2000) Biotic Invasions : Causes , Epidemiology , Global Consequences , and Control. Ecological Applications, 10, 689-710. doi: https://doi.org/10.1890/1051-0761(2000)010[0689:BICEGC]2.0.CO;2 Mallez, S., Castagnone, C., Lombaert, E., Castagnone-Sereno, P. and Guillemaud, T. (2021) Inference of the worldwide invasion routes of the pinewood nematode Bursaphelenchus xylophilus using approximate Bayesian computation analysis. bioRxiv, 452326, ver. 6 peer-reviewed and recommended by Peer community in Zoology. doi: https://doi.org/10.1101/452326 | Inference of the worldwide invasion routes of the pinewood nematode Bursaphelenchus xylophilus using approximate Bayesian computation analysis | Sophie Mallez, Chantal Castagnone, Eric Lombaert, Philippe Castagnone-Sereno, Thomas Guillemaud | <p>Population genetics have been greatly beneficial to improve knowledge about biological invasions. Model-based genetic inference methods, such as approximate Bayesian computation (ABC), have brought this improvement to a higher level and are now... | | Biogeography, Biological invasions, Ecology, Evolution, Genetics/Genomics, Herbivores, Invertebrates, Molecular biology, Nematology | Stéphane Dupas | 2020-09-15 10:59:41 | ||
24 Jun 2022

Dopamine pathway characterization during the reproductive mode switch in the pea aphidIn search of the links between environmental signals and polyphenismRecommended by Mathieu Joron based on reviews by Antonia Monteiro and 2 anonymous reviewersPolyphenisms offer an opportunity to study the links between phenotype, development, and environment in a controlled genomic context (Simpson, Sword, & Lo, 2011). In organisms with short generation times, individuals living and developing in different seasons encounter different environmental conditions. Adaptive plasticity allows them to express different phenotypes in response to seasonal cues, such as temperature or photoperiod. Such phenotypes can be morphological variants, for instance displaying different wing patterns as seen in butterflies (Brakefield & Larsen, 1984; Nijhout, 1991; Windig, 1999), or physiological variants, characterized for instance by direct development vs winter diapause in temperate insects (Dalin & Nylin, 2012; Lindestad, Wheat, Nylin, & Gotthard, 2019; Shearer et al., 2016). Many aphids display cyclical parthenogenesis, a remarkable seasonal polyphenism for reproductive mode (Tagu, Sabater-Muñoz, & Simon, 2005), also sometimes coupled with wing polyphenism (Braendle, Friebe, Caillaud, & Stern, 2005), which allows them to switch between parthenogenesis during spring and summer to sexual reproduction and the production of diapausing eggs before winter. In the pea aphid Acyrthosiphon pisum, photoperiod shortening results in the production, by parthenogenetic females, of embryos developing into the parthenogenetic mothers of sexual individuals. The link between parthenogenetic reproduction and sexual reproduction, therefore, occurs over two generations, changing from a parthenogenetic form producing parthenogenetic females (virginoparae), to a parthenogenetic form producing sexual offspring (sexuparae), and finally sexual forms producing overwintering eggs (Le Trionnaire et al., 2022). The molecular basis for the transduction of the environmental signal into reproductive changes is still unknown, but the dopamine pathway is an interesting candidate. Form-specific expression of certain genes in the dopamine pathway occurs downstream of the perception of the seasonal cue, notably with a marked decrease in the heads of embryos reared under short-day conditions and destined to become sexuparae. Dopamine has multiple roles during development, with one mode of action in cuticle melanization and sclerotization, and a neurological role as a synaptic neurotransmitter. Both modes of action might be envisioned to contribute functionally to the perception and transduction of environmental signals. In this study, Le Trionnaire and colleagues aim at clarifying this role in the pea aphid (Le Trionnaire et al., 2022). Using quantitative RT-PCR, RNA-seq, and in situ hybridization of RNA probes, they surveyed the timing and spatial patterns of expression of dopamine pathway genes during the development of different stages of embryo to larvae reared under long and short-day conditions, and destined to become virginoparae or sexuparae females, respectively. The genes involved in the synaptic release of dopamine generally did not show differences in expression between photoperiodic treatments. By contrast, pale and ddc, two genes acting upstream of dopamine production, generally tended to show a downregulation in sexuparare embryo, as well as genes involved in cuticle development and interacting with the dopamine pathway. The downregulation of dopamine pathway genes observed in the heads of sexuparare juveniles is already detectable at the embryonic stage, suggesting embryos might be sensing environmental cues leading them to differentiate into sexuparae females. The way pale and ddc expression differences could influence environmental sensitivity is still unclear. The results suggest that a cuticle phenotype specifically in the heads of larvae could be explored, perhaps in the form of a reduction in cuticle sclerotization and melanization which might allow photoperiod shortening to be perceived and act on development. Although its causality might be either way, such a link would be exciting to investigate, yet the existence of cuticle differences between the two reproductive types is still a hypothesis to be tested. The lack of differences in the expression of synaptic release genes for dopamine might seem to indicate that the plastic response to photoperiod is not mediated via neurological roles. Yet, this does not rule out the role of decreasing levels of dopamine in mediating this response in the central nervous system of embryos, even if the genes regulating synaptic release are equally expressed. To test for a direct role of ddc in regulating the reproductive fate of embryos, the authors have generated CrispR-Cas9 knockout mutants. Those mutants displayed egg cuticle melanization, but with lethal effects, precluding testing the effect of ddc at later stages in development. Gene manipulation becomes feasible in the pea aphid, opening up certain avenues for understanding the roles of other genes during development. This study adds nicely to our understanding of the intricate changes in gene expression involved in polyphenism. But it also shows the complexity of deciphering the links between environmental perception and changes in physiology, which mobilise multiple interacting gene networks. In the era of manipulative genetics, this study also stresses the importance of understanding the traits and phenotypes affected by individual genes, which now seems essential to piece the puzzle together. References Braendle C, Friebe I, Caillaud MC, Stern DL (2005) Genetic variation for an aphid wing polyphenism is genetically linked to a naturally occurring wing polymorphism. Proceedings of the Royal Society B: Biological Sciences, 272, 657–664. https://doi.org/10.1098/rspb.2004.2995 Brakefield PM, Larsen TB (1984) The evolutionary significance of dry and wet season forms in some tropical butterflies. Biological Journal of the Linnean Society, 22, 1–12. https://doi.org/10.1111/j.1095-8312.1984.tb00795.x Dalin P, Nylin S (2012) Host-plant quality adaptively affects the diapause threshold: evidence from leaf beetles in willow plantations. Ecological Entomology, 37, 490–499. https://doi.org/10.1111/j.1365-2311.2012.01387.x Le Trionnaire G, Hudaverdian S, Richard G, Tanguy S, Gleonnec F, Prunier-Leterme N, Gauthier J-P, Tagu D (2022) Dopamine pathway characterization during the reproductive mode switch in the pea aphid. bioRxiv, 2020.03.10.984989, ver. 4 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.1101/2020.03.10.984989 Lindestad O, Wheat CW, Nylin S, Gotthard K (2019) Local adaptation of photoperiodic plasticity maintains life cycle variation within latitudes in a butterfly. Ecology, 100, e02550. https://doi.org/10.1002/ecy.2550 Nijhout HF (1991). The development and evolution of butterfly wing patterns. Washington, DC: Smithsonian Institution Press. Shearer PW, West JD, Walton VM, Brown PH, Svetec N, Chiu JC (2016) Seasonal cues induce phenotypic plasticity of Drosophila suzukii to enhance winter survival. BMC Ecology, 16, 11. https://doi.org/10.1186/s12898-016-0070-3 Simpson SJ, Sword GA, Lo N (2011) Polyphenism in Insects. Current Biology, 21, R738–R749. https://doi.org/10.1016/j.cub.2011.06.006 Tagu D, Sabater-Muñoz B, Simon J-C (2005) Deciphering reproductive polyphenism in aphids. Invertebrate Reproduction & Development, 48, 71–80. https://doi.org/10.1080/07924259.2005.9652172 Windig JJ (1999) Trade-offs between melanization, development time and adult size in Inachis io and Araschnia levana (Lepidoptera: Nymphalidae)? Heredity, 82, 57–68. https://doi.org/10.1038/sj.hdy.6884510 | Dopamine pathway characterization during the reproductive mode switch in the pea aphid | Gaël Le Trionnaire, Sylvie Hudaverdian, Gautier Richard, Sylvie Tanguy, Florence Gleonnec, Nathalie Prunier-Leterme, Jean-Pierre Gauthier, Denis Tagu | <p>Aphids are major pests of most of the crops worldwide. Such a success is largely explained by the remarkable plasticity of their reproductive mode. They reproduce efficiently by viviparous parthenogenesis during spring and summer generating imp... | | Development, Genetics/Genomics, Insecta, Molecular biology | Mathieu Joron | 2020-03-13 13:01:44 | ||
19 Jul 2024

Museomics of Carabus giant ground beetles shows an Oligocene origin and in situ Alpine diversificationNatural history collections continue to inform ground beetle genetics.Recommended by Felix Sperling based on reviews by Michael Caterino, Julian Dupuis and 1 anonymous reviewerSome of the biodiversity of our planet now exists only in museums, due to continuing habitat destruction and climate change. With more than 380 million entomological specimens already preserved in museums (Johnson and Owens 2023), there is much work left to document what we already have. Fortunately, new advances in DNA sequencing have given us the opportunity to get enormous amounts of information from dried specimens on pins. Johnson KR, Owens, (IFP. 2023) A global approach for natural history museum collections. Science 379,1192-1194(2023). https://doi.org/10.1126/science.adf6434 Pauli MT, Gauthier J, Labédan M, Blanc M, Bilat J, Toussaint EFA (2024) Museomics of Carabus giant ground beetles shows an Oligocene origin and in situ alpine diversification. bioRxiv, ver. 5 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.1101/2024.03.21.586057 Schmid, S., Genevest, R., Gobet, E., Suchan, T., Sperisen, C., Tinner, W. and Alvarez, N. (2017), HyRAD-X, a versatile method combining exome capture and RAD sequencing to extract genomic information from ancient DNA. Methods Ecol Evol, 8: 1374-1388. https://doi.org/10.1111/2041-210X.12785 Toussaint EFA, Gauthier J, Bilat J, Gillett CPDT, Gough HM, Lundkvist H, Blanc M, Muñoz-Ramírez CP, Alvarez N (2021) HyRAD-X Exome Capture Museomics Unravels Giant Ground Beetle Evolution, Genome Biology and Evolution, Volume 13, Issue 7, evab112, https://doi.org/10.1093/gbe/evab112 | Museomics of *Carabus* giant ground beetles shows an Oligocene origin and *in situ* Alpine diversification | Marie T. PAULI, Jeremy GAUTHIER, Marjorie LABEDAN, Mickael BLANC, Julia BILAT, Emmanuel F.A. TOUSSAINT | <p style="text-align: justify;">The development of museomics represents a major paradigm shift in the use of natural history collection specimens for systematics and evolutionary biology. New approaches in this field allow the sequencing of hundre... | | Insecta, Phylogeny, Systematics | Felix Sperling | 2024-03-27 15:30:31 | ||
26 Aug 2022
Within and among population differences in cuticular hydrocarbons in the seabird tick Ixodes uriaeSeabird tick diversification and cuticular hydrocarbonsRecommended by Felix Sperling based on reviews by 2 anonymous reviewersTicks are notorious vectors of diseases in humans and other vertebrates. Much effort has been expended to understand tick diversity and ecology with the aim of managing their populations to alleviate the misery they bring. Further, the fundamental question of whether ticks are usually host generalists or host specialists has been debated at length and is important both for understanding the mechanisms of their diversification as well as for focusing control of ticks [1]. One elegant resolution of this question is to consider most tick species to be global generalists but local specialists [1]. This is well illustrated in a series of studies of the seabird tick, Ixodes uriae, which is comprised of host-specific races that show genetic [2], morphological [3] and host performance [4] differences associated with the seabirds they feed on. Such a pattern has clear ramifications for sympatric speciation; however, the factors that potentially act to drive these differences have remained elusive. Dupraz et al. [5] have now made intriguing and important steps toward bridging the gap between demonstrating local patterns of tick host association and understanding the physiological mechanisms that may facilitate such divergences. They collected I. uriae ticks from the nests of two seabirds – Atlantic puffins and common guillemots – on the north side of Iceland. Four populations of ticks were sampled, with one island providing both puffin ticks and guillemot ticks, to give two tick populations from each of the two seabird host species. They then washed the ticks in solvent and analyzed the dissolved cuticular hydrocarbons (CHCs) using GC mass spectrometry, revealing 22 different hydrocarbon compounds common to most of these samples. CHCs are known to be important across arthropods for a variety of functions ranging from reducing water loss to facilitating communication and recognition between individuals with species. Dupraz et al. [5] found three hydrocarbons that distinguished puffin ticks most consistently from guillemot ticks. A cross-validation test for host type also assigned 75% of the tick pools to the seabird host of origin. However, with these limited sample sizes, statistical analysis revealed no significant difference in CHC profiles between the host types, although a tendency was evident. Nonetheless, this study revealed a number of potentially diagnostic CHCs for tick host type, as well as some that may be more diagnostic of locations. This provides a fascinating and actionable foundation for further work using additional sites and host types, as well as an entry point into discerning the mechanisms at play in producing the diversity, complexity and adaptability that make ticks such medical menaces. References | Within and among population differences in cuticular hydrocarbons in the seabird tick Ixodes uriae | Marlène Dupraz, Chloe Leroy, Thorkell Lindberg Thórarinsson, Patrizia d’Ettorre, Karen D. McCoy | <p>The hydrophobic layer of the arthropod cuticle acts to maintain water balance, but can also serve to transmit chemical signals via cuticular hydrocarbons (CHC), essential mediators of arthropod behavior. CHC signatures typically vary qualitativ... | | Acari, Biology, Ecology, Evolution | Felix Sperling | 2022-02-08 13:00:52 | ||
25 Mar 2022

Pre- and post-oviposition behavioural strategies to protect eggs against extreme winter cold in an insect with maternal careNew insights into maternal egg care in insects: egg transport as an adaptive behavior to extreme temperatures in the European earwigRecommended by Anna Cohuet based on reviews by Ana Rivero, Nicolas Sauvion and Wolf U. BlanckenhornBecause of the inability of eggs to move, the fitness of oviparous organisms is particularly dependent on the oviposition site. The choice of oviposition site by mothers is therefore the result of trade-offs between exposure to risk factors or favorable conditions such as the presence/absence of predators, the threat of extreme temperatures, the risk of desiccation, the presence and quality of nutritional resources... In addition to these trade-offs between different biotic and abiotic factors that determine oviposition site selection, the ability of mothers to move their eggs after oviposition is a game-changer in insect strategies to optimize egg development and survival [1]. Oviposition site selection combined with egg transport has been explored in insects in relation to the risk of exposure to egg parasitoids [2] or needs for oxygenation [3] but surprisingly has not been investigated in regards to temperatures. Considering egg transport in the ability of insects to adapt their behavior to environmental conditions and in particular to potential extreme temperatures is yet inherent in providing a complete picture of the diversity of behaviors that shape adaptation to temperature and potential tolerance to climate change. In this sense, the study presented by Tourneur et al. [4], explores whether insects capable of egg-care might use egg transport as an adaptive behavior to protect them from suboptimal or extreme temperatures. The study was conducted in the European earwig, Forficula auricularia Linnaeus, 1758, which is known to practice egg-care in a variety of ways, that presumably includes egg-transportation, for several weeks or months during winter until hatching. The authors characterized different life-history traits related to egg-laying, egg-transport, and egg-development in two device systems with three experimental temperature regimes in two populations of European earwigs from Canada. The inclusion of two populations, which turned out to belong to two clades, allowed the identification of a diversity of behaviors although this did not allow to attribute the differences between the two populations to specific population differences, genetic differences, or to their geographical origins. Interestingly, the study showed that oviposition site selection in the European earwig is driven by temperature and that in winter temperatures, female earwigs may move their eggs to warmer temperatures that are adequate for hatching. These results are original in the sense that they highlight new adaptive strategies in female insects used during the post-oviposition stage to protect their eggs from temperature changes. In the current context of climate change and potential changes in selective pressures, the study contributes to the understanding of the wide range of strategies deployed by insects to adapt to the temperature. This appears essential to predict and anticipate the consequences of global instability, it also describes from an academic point of view a new and fascinating adaptive strategy in an overlooked biological system. References [1] Machado G, Trumbo ST (2018) Parental care. In: Insect Behavior, pp. 203–218. Oxford University Press, Oxford. https://doi.org/10.1093/oso/9780198797500.003.0014 [2] Carrasco D, Kaitala A (2009) Egg-laying tactic in Phyllomorpha laciniata in the presence of parasitoids. Entomologia Experimentalis et Applicata, 131, 300–307. https://doi.org/10.1111/j.1570-7458.2009.00857.x [3] Smith RL (1997) Evolution of paternal care in the giant water bugs (Heteroptera: Belostomatidae). In: The Evolution of Social Behaviour in Insects and Arachnids (eds Crespi BJ, Choe JC), pp. 116–149. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511721953.007 [4] Tourneur J-C, Cole C, Vickruck J, Dupont S, Meunier J (2022) Pre- and post-oviposition behavioural strategies to protect eggs against extreme winter cold in an insect with maternal care. bioRxiv, 2021.11.23.469705, ver. 3 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.1101/2021.11.23.469705 | Pre- and post-oviposition behavioural strategies to protect eggs against extreme winter cold in an insect with maternal care | Jean-Claude Tourneur, Claire Cole, Jess Vickruck, Simon Dupont, Joel Meunier | <p style="text-align: justify;">Depositing eggs in an area with adequate temperature is often crucial for mothers and their offspring, as the eggs are immobile and therefore cannot avoid exposure to sub-optimal temperatures. However, the importanc... | | Behavior, Ecology, Evolution, Insecta, Invertebrates, Life histories | Anna Cohuet | 2021-11-24 16:43:06 | ||
25 Aug 2021
Up and to the light: intra- and interspecific variability of photo- and geo-tactic oviposition preferences in genus TrichogrammaNew insights into oviposition preference of 5 Trichogramma speciesRecommended by Joël Meunier based on reviews by Kévin Tougeron and Eveline C. Verhulst
Insects exhibit a great diversity of life-history traits that often vary not only between species but also between populations of the same species (Flatt and Heyland, 2011). A better understanding of the variation in these traits can be of paramount importance when it comes to species of economic and agricultural interest (Wilby and Thomas, 2002). In particular, the control of the development and expansion of agricultural pests generally requires a good understanding of the parameters that favour the reproduction of these pests and/or the reproduction of the species used to control them (Bianchi et al., 2013; Gäde and Goldsworthy, 2003). Parasitoid wasps of the genus Trichogramma are a classic example of insects involved in pest control (Smith, 1996). This genus comprises over 200 species worldwide, which have been used to control populations of a wide range of lepidopteran pests since the 1900s (Flanders, 1930; Hassan, 1993). Despite its common use, the egg-laying preference of this genus is only partially known. For example, all Trichogramma species are often thought to have positive phototaxis (or negative geotaxis) (e.g. Brower & Cline, 1984; van Atta et al., 2015), but comprehensive studies simultaneously testing this (or other) parameter among Trichogramma species and populations remain rare. This is exactly the aim of the present study (Burte et al., 2021). Using a new experimental approach based on automatic image analysis, the authors compared the photo- and geo-tactic oviposition preference among 5 Trichogramma species from 25 populations. Their results first confirm that most Trichogramma species and populations prefer light to shade, and higher to lower positions for oviposition. Interestingly, they also reveal that the levels of preference for light and gravity show inter- and intraspecific variation (probably due to local adaptation to different strata) and that both preferences tend to relax over time. Overall, this study provides important information for improving the use of Trichogramma species as biological agents. For example, it may help to establish breeding lines adapted to the microhabitat and/or growing parts of plants on which agricultural pests lay eggs most. Similarly, it suggests that the use of multiple strains with different microhabitat selection preferences could lead to better coverage of host plants, as well as a reduction in intraspecific competition in the preferred parts. Finally, this study provides a new methodology to efficiently and automatically study oviposition preferences in Trichogramma, which could be used in other insects with a particularly small size. References Bianchi, F. J. J. A., Schellhorn, N. A. and Cunningham, S. A. (2013). Habitat functionality for the ecosystem service of pest control: reproduction and feeding sites of pests and natural enemies. Agricultural and Forest Entomology, 15, 12–23. https://doi.org/10.1111/j.1461-9563.2012.00586.x Burte V., Perez G., Ayed F., Groussier G., Mailleret L, van Oudenhove L. and Calcagno V. (2021). Up and to the light: intra- and interspecific variability of photo-and geo-tactic oviposition preferences in genus Trichogramma. bioRxiv, 2021.03.30.437671, ver. 4 peer-reviewed and recommended by PCI Zoology. https://doi.org/10.1101/2021.03.30.437671 Brower, J. H. and Cline, L. D. (1984). Response of Trichogramma pretiosum and T. evanescens to Whitelight, Blacklight or NoLight Suction Traps. The Florida Entomologist, 67, 262–268. https://doi.org/10.2307/3493947 Flanders, S. E. (1930). Mass production of egg parasites of the genus Trichogramma. Hilgardia, 4, 465–501. https://doi.org/10.3733/hilg.v04n16p465 Flatt, T. and Heyland, A. (2011). Mechanisms of life history evolution: the genetics and physiology of life history traits and trade-offs. Oxford University Press. https://doi.org/10.1093/acprof:oso/9780199568765.001.0001 Gäde, G. and Goldsworthy, G. J. (2003). Insect peptide hormones: a selective review of their physiology and potential application for pest control. Pest Management Science, 59, 1063–1075. https://doi.org/10.1002/ps.755 Hassan, S. A. (1993). The mass rearing and utilization of Trichogramma to control lepidopterous pests: Achievements and outlook. Pesticide Science, 37, 387–391. https://doi.org/10.1002/ps.2780370412 Smith, S. M. (1996). Biological Control with Trichogramma : Advances, Successes, and Potential of Their Use. Annual Review of Entomology, 41, 375–406. https://doi.org/10.1146/annurev.en.41.010196.002111 van Atta, K. J., Potter, K. A. and Woods, H. A. (2015). Effects of UV-B on Environmental Preference and Egg Parasitization by Trichogramma Wasps (Hymenoptera: Trichogrammatidae). Journal of Entomological Science, 50, 318–325. https://doi.org/10.18474/JES15-09.1 Wilby, A. and Thomas, M. B. (2002). Natural enemy diversity and pest control: patterns of pest emergence with agricultural intensification. Ecology Letters, 5, 353–360. https://doi.org/10.1046/j.1461-0248.2002.00331.x | Up and to the light: intra- and interspecific variability of photo- and geo-tactic oviposition preferences in genus Trichogramma | Burte, V., Perez, G., Ayed, F. , Groussier, G., Mailleret, L., van Oudenhove, L. and Calcagno, V. | <p>Trichogramma are parasitic microwasps much used as biological control agents. The genus is known to harbor tremendous diversity, at both inter- and intra-specific levels. The successful selection of Trichogramma strains for biocontrol depends o... | | Behavior, Biocontrol, Biodiversity, Ecology, Insecta, Parasitology, Pest management, Systematics, Terrestrial | Joël Meunier | Kévin Tougeron, Eveline C. Verhulst | 2021-04-02 16:10:28 | |
21 Jun 2023
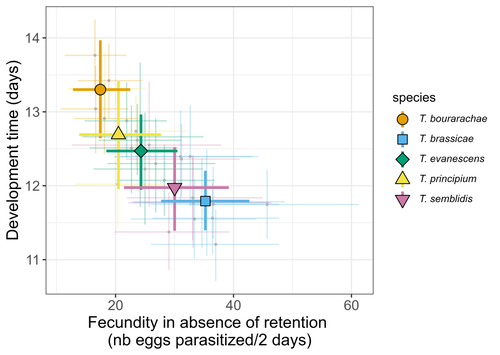
Life-history traits, pace of life and dispersal among and within five species of Trichogramma wasps: a comparative analysisThe relationship between dispersal and pace-of-life at different scalesRecommended by Jacques Deere based on reviews by Mélanie Thierry and 1 anonymous reviewerThe sorting of organisms along a fast-slow continuum through correlations between life history traits is a long-standing framework (Stearns 1983) and corresponds to the pace-of-life axis. This axis represents the variation in a continuum of life-history strategies, from fast-reproducing short-lived species to slow-reproducing long-lived species. The pace-of-life axis has been the focus of much research largely in mammals, birds, reptiles and plants but less so in invertebrates (Salguero-Gómez et al. 2016; Araya-Ajoy et al. 2018; Healy et al. 2019; Bakewell et al. 2020). Outcomes from this research have highlighted variation across taxa on this axis and mixed support for, and against, patterns expected of the pace-of-life continuum. Given this, a greater understanding of the variation of the pace-of-life across-, and within, taxa are needed. Indeed, Guicharnard et al. (2023) highlight several points regarding our broader understanding of pace-of-life. In general, invertebrates are poorly represented, the variation of pace-of-life across taxonomic scales is less well understood and the relationship between pace-of-life and dispersal, a key life history, requires more attention. Here, Guicharnard et al. (2023) provide a first attempt at addressing the relationship between dispersal and pace-of-life at different scales. The authors, under controlled conditions, investigated how life-history traits and effective dispersal covary for 28 lines from five species of endoparasitoid wasps from the genus Trichogramma. At the species level negative correlations were found between development time and fecundity, matching pace-of-life axis predictions. Although this correlation was not found to be significant among lines, within species, a similar pattern of a negative correlation was observed. This outcome matches previous findings that consistent pace-of-life axes become more difficult to find at lower taxonomic levels. Unlike the other life-history traits measured, effective dispersal showed no evidence of differences between species or between lines. The authors also found no correlation between effective dispersal and other-life history traits which suggests no dispersal/life-history syndromes in the species investigated. One aspect that was not assessed was the impact of density dependence on pace-of-life and effective dispersal, largely as this was a first step in assessing relationship of dispersal with pace-of-life at different scales. However, the authors do acknowledge the importance of future studies incorporating density dependence and that such studies could potentially lead to more generalizable understanding of pace-of-life and dispersal within Trichogramma. A pleasant addition was the link to potential implications for biocontrol. This addition showed an awareness by the authors of how insights into pace-of-life can have an applied component. The results of the study highlighted that selecting for specific lines of a species, to maximise a trait of interest at the cost of another, may not be as effective as selecting different species when implementing biocontrol. This is especially important as often single, established species used in biocontrol are favoured without consideration of the potential of other species which can lead to more efficient biocontrol. REFERENCES Araya-Ajoy, Y.G., Bolstad, G.H., Brommer, J., Careau, V., Dingemanse, N.J. & Wright, J. (2018). Demographic measures of an individual's "pace of life": fecundity rate, lifespan, generation time, or a composite variable? Behavioral Ecology and Sociobiology, 72, 75. | Life-history traits, pace of life and dispersal among and within five species of *Trichogramma* wasps: a comparative analysis | Chloé Guicharnaud, Géraldine Groussier, Erwan Beranger, Laurent Lamy, Elodie Vercken, Maxime Dahirel | <p>Major traits defining the life history of organisms are often not independent from each other, with most of their variation aligning along key axes such as the pace-of-life axis. We can define a pace-of-life axis structuring reproduction and de... | | Biology, Ecology, Insecta, Invertebrates, Life histories | Jacques Deere | 2023-01-25 18:15:20 | ||
14 Nov 2023

Time-course of antipredator behavioral changes induced by the helminth Pomphorhynchus laevis in its intermediate host Gammarus pulex: the switch in manipulation according to parasite developmental stage differs between behaviorsExploring manipulative strategies of a trophically-transmitted parasite across its ontogenyRecommended by Thierry Lefevre based on reviews by Adèle Mennerat and 1 anonymous reviewerThe intricate relationships between parasites and their hosts often involve a choreography of behavioral changes, with parasites manipulating their hosts in a way that enhances - or seemingly enhances – their transmission (Hughes et al., 2012; Moore, 2002; Poulin, 2010). Host manipulation is increasingly acknowledged as a pervasive adaptive transmission strategy employed by parasites, and as such is one of the most remarkable manifestations of the extended phenotype (Dawkins, 1982). In this laboratory study, Rigaud et al. (2023) delved into the time course of antipredator behavioral modifications induced by the acanthocephalan Pomphorhynchus laevis in its amphipod intermediate host Gammarus pulex. This system has a good foundation of prior knowledge (Bakker et al., 2017; Fayard et al., 2020; Perrot-Minnot et al., 2023), nicely drawn upon for the present work. This parasite orchestrates a switch from predation suppression, during the noninfective phase, to predation enhancement upon maturation. Specifically, G. pulex infected with the non-infective acanthella stage of the parasite can exhibit increased refuge use and reduced activity compared to uninfected individuals (Dianne et al., 2011, 2014), leading to decreased predation by trout (Dianne et al., 2011). In contrast, upon reaching the infective cystacanth stage, the parasite can enhance the susceptibility of its host to trout predation (Dianne et al., 2011). The present work aimed to understand the temporal sequence of these behavioral changes across the entire ontogeny of the parasite. The results confirmed the protective role of P. laevis during the acanthella stage, wherein infected amphipods exhibited heightened refuge use. This protective manipulation, however, became significant only later in the parasite's ontogeny, suggesting a delayed investment strategy, possibly influenced by the extended developmental time of P. laevis. The protective component wanes upon reaching the cystacanth stage, transitioning into an exposure strategy, aligning with theoretical predictions and previous empirical work (Dianne et al., 2011; Parker et al., 2009). The switch was behavior-specific. Unlike the protective behavior, a decline in the amphipod activity rate manifested early in the acanthella stage and persisted throughout development, suggesting potential benefits of reduced activity for the parasite across multiple stages. Furthermore, the findings challenge previous assumptions regarding the condition-dependency of manipulation, revealing that the parasite-induced behavioral changes predominantly occurred in the presence of cues signaling potential predators. Finally, while amphipods infected with acanthella stages displayed survival rates comparable to their uninfected counterparts, increased mortality was observed in those infected with cystacanth stages. Understanding the temporal sequence of host behavioral changes is crucial for deciphering whether it is adaptive to the parasite or not. This study stands out for its meticulous examination of multiple behaviors over the entire ontogeny of the parasite highlighting the complexity and condition-dependent nature of manipulation. The protective-then-expose strategy emerges as a dynamic process, finely tuned to the developmental stages of the parasite and the ecological challenges faced by the host. The delayed emergence of protective behaviors suggests a strategic investment by the parasite, with implications for the host's survival and the parasite's transmission success. The differential impact of infection on refuge use and activity rate further emphasizes the need for a multidimensional approach in studying parasitic manipulation (Fayard et al., 2020). This complexity demands further exploration, particularly in deciphering how trophically-transmitted parasites shape the behavioral landscape of their intermediate hosts and its temporal dynamic (Herbison, 2017; Perrot-Minnot & Cézilly, 2013). As we discover the many subtleties of these parasitic manipulations, new avenues of research are unfolding, promising a deeper understanding of the ecology and evolution of host-parasite interactions. References Bakker, T. C. M., Frommen, J. G., & Thünken, T. (2017). Adaptive parasitic manipulation as exemplified by acanthocephalans. Ethology, 123(11), 779–784. https://doi.org/10.1111/eth.12660 Dawkins, R. (1982). The extended phenotype: The long reach of the gene (Reprinted). Oxford University Press. Dianne, L., Perrot-Minnot, M.-J., Bauer, A., Gaillard, M., Léger, E., & Rigaud, T. (2011). Protection first then facilitation: A manipulative parasite modulates the vulnerability to predation of its intermediate host according to its own developmental stage. Evolution, 65(9), 2692–2698. https://doi.org/10.1111/j.1558-5646.2011.01330.x Dianne, L., Perrot-Minnot, M.-J., Bauer, A., Guvenatam, A., & Rigaud, T. (2014). Parasite-induced alteration of plastic response to predation threat: Increased refuge use but lower food intake in Gammarus pulex infected with the acanothocephalan Pomphorhynchus laevis. International Journal for Parasitology, 44(3–4), 211–216. https://doi.org/10.1016/j.ijpara.2013.11.001 Fayard, M., Dechaume‐Moncharmont, F., Wattier, R., & Perrot‐Minnot, M. (2020). Magnitude and direction of parasite‐induced phenotypic alterations: A meta‐analysis in acanthocephalans. Biological Reviews, 95(5), 1233–1251. https://doi.org/10.1111/brv.12606 Herbison, R. E. H. (2017). Lessons in Mind Control: Trends in Research on the Molecular Mechanisms behind Parasite-Host Behavioral Manipulation. Frontiers in Ecology and Evolution, 5, 102. https://doi.org/10.3389/fevo.2017.00102 Hughes, D. P., Brodeur, J., & Thomas, F. (2012). Host manipulation by parasites. Oxford university press. Moore, J. (2002). Parasites and the behavior of animals. Oxford University Press. Parker, G. A., Ball, M. A., Chubb, J. C., Hammerschmidt, K., & Milinski, M. (2009). When should a trophically transmitted parasite manipulate its host? Evolution, 63(2), 448–458. https://doi.org/10.1111/j.1558-5646.2008.00565.x Perrot-Minnot, M.-J., & Cézilly, F. (2013). Investigating candidate neuromodulatory systems underlying parasitic manipulation: Concepts, limitations and prospects. Journal of Experimental Biology, 216(1), 134–141. https://doi.org/10.1242/jeb.074146 Perrot-Minnot, M.-J., Cozzarolo, C.-S., Amin, O., Barčák, D., Bauer, A., Filipović Marijić, V., García-Varela, M., Servando Hernández-Orts, J., Yen Le, T. T., Nachev, M., Orosová, M., Rigaud, T., Šariri, S., Wattier, R., Reyda, F., & Sures, B. (2023). Hooking the scientific community on thorny-headed worms: Interesting and exciting facts, knowledge gaps and perspectives for research directions on Acanthocephala. Parasite, 30, 23. https://doi.org/10.1051/parasite/2023026 Poulin, R. (2010). Parasite Manipulation of Host Behavior. In Advances in the Study of Behavior (Vol. 41, pp. 151–186). Elsevier. https://doi.org/10.1016/S0065-3454(10)41005-0 Rigaud, T., Balourdet, A., & Bauer, A. (2023). Time-course of antipredator behavioral changes induced by the helminth Pomphorhynchus laevis in its intermediate host Gammarus pulex: The switch in manipulation according to parasite developmental stage differs between behaviors. bioRxiv, ver. 6 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.1101/2023.04.25.538244 | Time-course of antipredator behavioral changes induced by the helminth *Pomphorhynchus laevis* in its intermediate host *Gammarus pulex*: the switch in manipulation according to parasite developmental stage differs between behaviors | Thierry Rigaud, Aude Balourdet, Alexandre Bauer | <p style="text-align: justify;">Many trophically transmitted parasites with complex life cycles manipulate their intermediate host antipredatory defenses in ways facilitating their transmission to final host by predation. Some parasites also prote... | | Aquatic, Behavior, Crustacea, Invertebrates, Parasitology | Thierry Lefevre | 2023-06-20 15:49:32 |




