
Latest recommendations

| Id | Title▼ | Authors | Abstract | Picture | Thematic fields | Recommender | Reviewers | Submission date | |
|---|---|---|---|---|---|---|---|---|---|
20 Dec 2022

Non-target effects of ten essential oils on the egg parasitoid Trichogramma evanescensSide effects of essential oils on pest natural enemiesRecommended by Cedric Pennetier based on reviews by Olivier Roux and 2 anonymous reviewersIntegrated pest management relies on the combined use of different practices in time and/or space. The main objectives are to better control pests, not to induce too much selective pressure on resistance mechanisms present in pest populations and to minimize non-targeted effects on the ecosystem [1]. The efficiency of such a strategy requires at least additional or synergistic effects of chosen tools against targeted pest population in a specific environment. Any antagonistic effect on targeted or non-targeted organisms might reduce control effort to nil even worst. Van Oudenhove et al [2] raised the question of the interaction between botanical pesticides (BPs) and egg parasitoids. Each of these two strategies used for pest management present advantages and are described as eco-friendly. First, the use of parasitoids is a great example of biological control and is massively used in a broad range of crop production in different ecological settings. Second, BPs, especially essential oils (EOs) used for a wide range of activities on pests (repellent, antifeedant, antiovipositant, ovicidal, larvicidal and simply pesticidal) present low-toxicity to non-target vertebrates and do not last too long in the environment. Combining these two strategies might be considered as a great opportunity to better pest control with minimized impact on environment. However, EOs used to target a wide range of pest might directly or indirectly affect parasitoids. Van Oudenhove et al [2] focused their study on non-target effects of 10 essentials oils with pesticide potential on larval development and egg-seeking behaviour of five strains of the biocontrol agent Trichogramma evanescens. Within two laboratory experiments mimicing EOs fumigation (i.e. contactless EOs exposure), the authors evaluated (1) the toxicity of EOs on parasitoid development and (2) the repellent effect of these EOs on adult wasps. They confirmed that contactless exposure of EOs can (1) induce mortality during pre-imaginal development (more acute at the pupal stage) and (2) induce behavioural avoidance of EOs odour plume. These experiments ran onto five strains of T. evanescens also highlighted the variation of the effects of EOs among parasitoid strains. The complex and dynamic interaction between pest, plant, parasitoid (a natural enemy) and their environment is disturbed by EOs. EOs plumes are also dynamic and variable upon the environmental conditions. The results of van Oudenhove et al. experimentally illustrate such a complexity by describing opposite effects (repellent and attractive) of the same EO on the behaviour of two T. evanescens strains. These contrasting results led us to question more broadly the non-target effects of pest management programs based on EOs fumigation on natural enemies. Finally, the limits of this experimental study as discussed in the paper draw research avenues taking into account biotic variables such as plant chemical cues, odour plume dynamics, individual behavioural experiences and abiotic variables such as temperature, light and gravity [3] in laboratory, semi-field and field experiments. Facing such a complexity, modelling studies at fine scale in time and space have the operational objective to help farmers to choose the best IPM strategy regarding their environment (as illustrated for aphid population management in the recent review by Stell et al. [4]). But before such research effort to be undertaken, Van Oudenhove et al study [2] sounds like an alert for a cautious use of EOs in pest control programs that integrate biological control with parasitoids.
References [1] Fauvergue, X. Biocontrôle Elements Pour Une Protection Agroecologique des Cultures; Éditions Quae: Versailles, France, 2020. [2] van Oudenhove L, Cazier A, Fillaud M, Lavoir AV, Fatnassi H, Pérez G, Calcagno V. Non-target effects of ten essential oils on the egg parasitoid Trichogramma evanescens. bioRxiv 2022.01.14.476310, ver. 4 peer-reviewed and recommended by PCI Zoology. https://doi.org/10.1101/2022.01.14.476310 [3] Victor Burte, Guy Perez, Faten Ayed, Géraldine Groussier, Ludovic Mailleret, Louise van Oudenhove and Vincent Calcagno (2022) Up and to the light: intra- and interspecific variability of photo- and geo-tactic oviposition preferences in genus Trichogramma, Peer Community Journal, 2: e3. https://doi.org/10.24072/pcjournal.78 [4] Stell E, Meiss H, Lasserre-Joulin F, Therond O. Towards Predictions of Interaction Dynamics between Cereal Aphids and Their Natural Enemies: A Review. Insects 2022, 13, 479. https://doi.org/10.3390/insects13050479 | Non-target effects of ten essential oils on the egg parasitoid Trichogramma evanescens | Louise van Oudenhove, Aurélie Cazier, Marine Fillaud, Anne-Violette Lavoir, Hicham Fatnassi, Guy Pérez, Vincent Calcagno | <p style="text-align: justify;">Essential oils (EOs) are increasingly used as biopesticides due to their insecticidal potential. This study addresses their non-target effects on a biological control agent: the egg parasitoid <em>Trichogramma evane... | | Behavior, Biochemistry, Biocontrol, Biodiversity, Computer modelling, Conservation biology, Demography/population dynamics, Development, Ecology, Insecta, Insectivores, Invertebrates, Life histories, Methodology, Pest management, Theoretical biolo... | Cedric Pennetier | 2022-01-31 16:05:32 | ||
26 Apr 2023
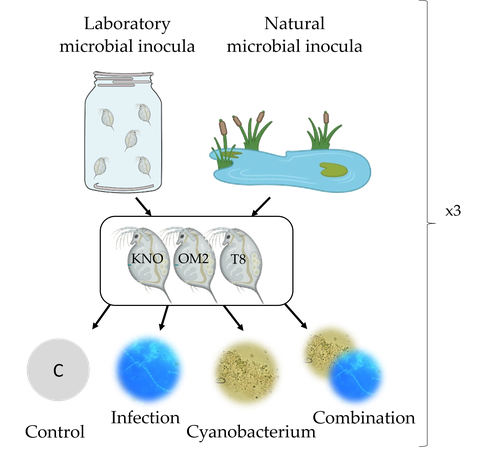
Microbiome mediated tolerance to biotic stressors: a case study of the interaction between a toxic cyanobacterium and an oomycete-like infection in Daphnia magnaMulti-stress responses depend on the microbiome in the planktonic crustacean DaphniaRecommended by Bertanne Visser and Mathilde Scheifler based on reviews by Natacha Kremer and 2 anonymous reviewers based on reviews by Natacha Kremer and 2 anonymous reviewers
The critical role that gut microbiota play in many aspects of an animal’s life, including pathogen resistance, detoxification, digestion, and nutritional physiology, is becoming more and more apparent (Engel and Moran 2013; Lindsay et al., 2020). Gut microbiota recruitment and maintenance can be largely affected by the surrounding environment (Chandler et al., 2011; Callens et al., 2020). The environment may thus dictate gut microbiota composition and diversity, which in turn can affect organismal responses to stress. Only few studies have, however, taken the gut microbiota into account to estimate life histories in response to multiple stressors in aquatic systems (Macke et al., 2016). Houwenhuyse et al., investigate how the microbiome affects life histories in response to ecologically relevant single and multiple biotic stressors (an oomycete-like parasite, and a toxic cyanobacterium) in Daphnia magna (Houwenhuyse et al., 2023). Daphnia is an excellent model, because this aquatic system lends itself extremely well for gut microbiota transplantation and manipulation. This is due to the possibility to sterilize eggs (making them free of bacteria), horizontal transmission of bacteria from the environment, and the relative ease of culturing genetically similar Daphnia clones in large numbers. The authors use an elegant experimental design to show that the Daphnia gut microbial community differs when derived from a laboratory versus natural inoculum, the latter being more diverse. The authors subsequently show that key life history traits (survival, fecundity, and body size) depend on the stressors (and combination thereof), the microbiota (structure and diversity), and Daphnia genotype. A key finding is that Daphnia exposed to both biotic stressors show an antagonistic interaction effect on survival (being higher), but only in individuals containing laboratory gut microbiota. The exact mechanism remains to be determined, but the authors propose several interesting hypotheses as to why Daphnia with more diverse gut microbiota do less well. This could be due, for example, to increased inter-microbe competition or an increased chance of contracting opportunistic, parasitic bacteria. For Daphnia with less diverse laboratory gut microbiota, a monopolizing species may be particularly beneficial for stress tolerance. Alongside these interesting findings, the paper also provides extensive information about the gut microbiota composition (available in the supplementary files), which is a very useful resource for other researchers. Overall, this study reveals that multiple, interacting factors affect the performance of Daphnia under stressful conditions. Of importance is that laboratory studies may be based on simpler microbiota systems, meaning that stress responses measured in the laboratory may not accurately reflect what is happening in nature. REFERENCES Callens M, De Meester L, Muylaert K, Mukherjee S, Decaestecker E. The bacterioplankton community composition and a host genotype dependent occurrence of taxa shape the Daphnia magna gut bacterial community. FEMS Microbiology Ecology. 2020;96(8):fiaa128. https://doi.org/10.1093/femsec/fiaa128 Chandler JA, Lang JM, Bhatnagar S, Eisen JA, Kopp A. Bacterial communities of diverse Drosophila species: ecological context of a host-microbe model system. PLOS Genetics. 2011;7(9):e1002272. https://doi.org/10.1371/journal.pgen.1002272 Engel P, Moran NA. The gut microbiota of insects - diversity in structure and function. FEMS Microbiology Reviews. 2013;37(5):699-735. https://doi.org/10.1111/1574-6976.12025 Houwenhuyse S, Bulteel L, Vanoverberghe I, Krzynowek A, Goel N et al. Microbiome mediated tolerance to biotic stressors: a case study of the interaction between a toxic cyanobacterium and an oomycete-like infection in Daphnia magna. 2023. OSF, ver. 2 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.31219/osf.io/9n4mg Lindsay EC, Metcalfe NB, Llewellyn MS. The potential role of the gut microbiota in shaping host energetics and metabolic rate. Journal of Animal Ecology. 2020;89(11):2415-2426. https://doi.org/10.1111/1365-2656.13327 Macke E, Tasiemski A, Massol F, Callens M, Decaestecker E. Life history and eco-evolutionary dynamics in light of the gut microbiota. Oikos. 2017;126(4):508-531. https://doi.org/10.1111/oik.03900 | Microbiome mediated tolerance to biotic stressors: a case study of the interaction between a toxic cyanobacterium and an oomycete-like infection in *Daphnia magna* | Shira Houwenhuyse*, Lore Bulteel*, Isabel Vanoverberghe, Anna Krzynowek, Naina Goel, Manon Coone, Silke Van den Wyngaert, Arne Sinnesael, Robby Stoks & Ellen Decaestecker | <p style="text-align: justify;">Organisms are increasingly facing multiple, potentially interacting stressors in natural populations. The ability of populations coping with combined stressors depends on their tolerance to individual stressors and ... | | Aquatic, Biology, Crustacea, Ecology, Life histories, Symbiosis | Bertanne Visser | 2021-05-17 16:18:18 | ||
21 Jun 2023
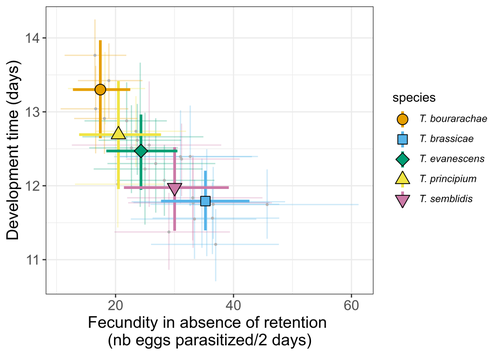
Life-history traits, pace of life and dispersal among and within five species of Trichogramma wasps: a comparative analysisThe relationship between dispersal and pace-of-life at different scalesRecommended by Jacques Deere based on reviews by Mélanie Thierry and 1 anonymous reviewerThe sorting of organisms along a fast-slow continuum through correlations between life history traits is a long-standing framework (Stearns 1983) and corresponds to the pace-of-life axis. This axis represents the variation in a continuum of life-history strategies, from fast-reproducing short-lived species to slow-reproducing long-lived species. The pace-of-life axis has been the focus of much research largely in mammals, birds, reptiles and plants but less so in invertebrates (Salguero-Gómez et al. 2016; Araya-Ajoy et al. 2018; Healy et al. 2019; Bakewell et al. 2020). Outcomes from this research have highlighted variation across taxa on this axis and mixed support for, and against, patterns expected of the pace-of-life continuum. Given this, a greater understanding of the variation of the pace-of-life across-, and within, taxa are needed. Indeed, Guicharnard et al. (2023) highlight several points regarding our broader understanding of pace-of-life. In general, invertebrates are poorly represented, the variation of pace-of-life across taxonomic scales is less well understood and the relationship between pace-of-life and dispersal, a key life history, requires more attention. Here, Guicharnard et al. (2023) provide a first attempt at addressing the relationship between dispersal and pace-of-life at different scales. The authors, under controlled conditions, investigated how life-history traits and effective dispersal covary for 28 lines from five species of endoparasitoid wasps from the genus Trichogramma. At the species level negative correlations were found between development time and fecundity, matching pace-of-life axis predictions. Although this correlation was not found to be significant among lines, within species, a similar pattern of a negative correlation was observed. This outcome matches previous findings that consistent pace-of-life axes become more difficult to find at lower taxonomic levels. Unlike the other life-history traits measured, effective dispersal showed no evidence of differences between species or between lines. The authors also found no correlation between effective dispersal and other-life history traits which suggests no dispersal/life-history syndromes in the species investigated. One aspect that was not assessed was the impact of density dependence on pace-of-life and effective dispersal, largely as this was a first step in assessing relationship of dispersal with pace-of-life at different scales. However, the authors do acknowledge the importance of future studies incorporating density dependence and that such studies could potentially lead to more generalizable understanding of pace-of-life and dispersal within Trichogramma. A pleasant addition was the link to potential implications for biocontrol. This addition showed an awareness by the authors of how insights into pace-of-life can have an applied component. The results of the study highlighted that selecting for specific lines of a species, to maximise a trait of interest at the cost of another, may not be as effective as selecting different species when implementing biocontrol. This is especially important as often single, established species used in biocontrol are favoured without consideration of the potential of other species which can lead to more efficient biocontrol. REFERENCES Araya-Ajoy, Y.G., Bolstad, G.H., Brommer, J., Careau, V., Dingemanse, N.J. & Wright, J. (2018). Demographic measures of an individual's "pace of life": fecundity rate, lifespan, generation time, or a composite variable? Behavioral Ecology and Sociobiology, 72, 75. | Life-history traits, pace of life and dispersal among and within five species of *Trichogramma* wasps: a comparative analysis | Chloé Guicharnaud, Géraldine Groussier, Erwan Beranger, Laurent Lamy, Elodie Vercken, Maxime Dahirel | <p>Major traits defining the life history of organisms are often not independent from each other, with most of their variation aligning along key axes such as the pace-of-life axis. We can define a pace-of-life axis structuring reproduction and de... | | Biology, Ecology, Insecta, Invertebrates, Life histories | Jacques Deere | 2023-01-25 18:15:20 | ||
28 Apr 2021
Inference of the worldwide invasion routes of the pinewood nematode Bursaphelenchus xylophilus using approximate Bayesian computation analysisExtracting the maximum historical information on pine wood nematode worldwide invasion from genetic dataRecommended by Stéphane Dupas based on reviews by Aude Gilabert and 1 anonymous reviewer
Redistribution of domesticated and non domesticated species by humans profoundly affected earth biogeography and in return human activities. This process accelerated exponentially since human expansion out of Africa, leading to the modern global, highly connected and homogenized, agriculture and trade system (Mack et al. 2000, Jaksic and Castro 2021), that threatens biological diversity and genetic resources. To accompany quarantine and control effort, the reconstruction of invasion routes provides valuable information that help identifying critical nodes and edges in the global networks (Estoup and Guillemaud 2010, Cristescu 2015). Historical records and genetic markers are the two major sources of information of this corpus of knowledge on Anthropocene historical phylogeography. With the advances of molecular genetics tools, the genealogy of these introductions events could be revisited and empowered. Due to their idiosyncrasy and intimate association with the contingency of human trades and activities, understanding the invasion and domestication routes require particular statistical tools (Fraimout et al. 2017). Because it encompasses all these theoretical, ecological and economical implications, I am pleased to recommend the readers of PCI Zoology this article by Mallez et al. (2021) on pine wood nematode invasion route inference from genetic markers using Approximate Bayesian Computation (ABC) methods. Economically and ecologically, this pest, is responsible for killing millions of pines worldwide each year. The results show these damages and the global genetic patterns are due to few events of successful introductions. The authors consider that this low probability of introductions success reinforces the idea that quarantine measures are efficient. This is illustrated in Europe where the pine-worm has been quarantined successfully in the Iberian Peninsula since 1999. Another relevant conclusion is that hybridization between invasive populations have not been observed and implied in the invasion process. Finally the present study reinforced the role of Asiatic bridgehead populations in invasion process including in Europe. Methodologically, for the first time, ABC was applied to this species. A total of 310 individual sequences were added to the Mallez et al. (2015) microsatellite dataset. Fraimoult et al. (2017) showed the interest to apply random forest to improve scenario selection in ABC framework. This method, implemented in the DiYABC software (Collin et al. 2020) for invasion route scenario selection allows to handle more complex scenario alternatives and was used in this study. In this article by Mallez et al. (2021), you will also find a clear illustration of the step-by-step approach to select scenario using ABC techniques (Lombaert et al. 2014). The rationale is to reduce number of scenario to be tested by assuming that most recent invasions cannot be the source of the most ancient invasions and to use posterior results on most ancient routes as prior hypothesis to distinguish following invasions. The other simplification is to perform classical population genetic analysis to characterize genetic units and representative populations prior to invasion routes scenarios selection by ABC. Yet, even when using the most advanced Bayesian inference methods, it is recognized by the authors that the method can be pushed to its statistical power limits. The method is appropriate when population show strong inter-population genetic structure. But the high number of differentiated populations in native area can be problematic since it is generally associated to incomplete sampling scheme. The hypothesis of ghost populations source allowed to bypass this difficulty, but the authors consider simulation studies are needed to assess the joint effect of genetic diversity and number of genetic markers on the inference results in such situation. Also the need to use a stepwise approach to reduce the number of scenario to test has to be considered with caution. Scenarios that are not selected but have non negligible posterior, cannot be ruled out in the constitution of next step scenarios hypotheses. Due to its interest to understand this major facet of Anthropocene, reconstruction of invasion routes should be more considered as a guide to damper biological homogenization process. References Collin, F.-D., Durif, G., Raynal, L., Lombaert, E., Gautier, M., Vitalis, R., Marin, J.-M. and Estoup, A. (2020) Extending Approximate Bayesian Computation with Supervised Machine Learning to infer demographic history from genetic polymorphisms using DIYABC Random Forest. Authorea. doi: https://doi.org/10.22541/au.159480722.26357192 Cristescu, M.E. (2015) Genetic reconstructions of invasion history. Molecular Ecology, 24, 2212–2225. doi: https://doi.org/10.1111/mec.13117 Estoup, A. and Guillemaud, T., (2010) Reconstructing routes of invasion using genetic data: Why, how and so what? Molecular Ecology, 9, 4113-4130. doi: https://doi.org/10.1111/j.1365-294X.2010.04773.x Fraimout, A., Debat, V., Fellous, S., Hufbauer, R.A., Foucaud, J., Pudlo, P., Marin, J.M., Price, D.K., Cattel, J., Chen, X., Deprá, M., Duyck, P.F., Guedot, C., Kenis, M., Kimura, M.T., Loeb, G., Loiseau, A., Martinez-Sañudo, I., Pascual, M., Richmond, M.P., Shearer, P., Singh, N., Tamura, K., Xuéreb, A., Zhang, J., Estoup, A. and Nielsen, R. (2017) Deciphering the routes of invasion of Drosophila suzukii by Means of ABC Random Forest. Molecular Biology and Evolution, 34, 980-996. doi: https://doi.org/10.1093/molbev/msx050 Jaksic, F.M. and Castro, S.A. (2021). Biological Invasions in the Anthropocene, in: Jaksic, F.M., Castro, S.A. (Eds.), Biological Invasions in the South American Anthropocene: Global Causes and Local Impacts. Springer International Publishing, Cham, pp. 19-47. doi: https://doi.org/10.1007/978-3-030-56379-0_2 Lombaert, E., Guillemaud, T., Lundgren, J., Koch, R., Facon, B., Grez, A., Loomans, A., Malausa, T., Nedved, O., Rhule, E., Staverlokk, A., Steenberg, T. and Estoup, A. (2014) Complementarity of statistical treatments to reconstruct worldwide routes of invasion: The case of the Asian ladybird Harmonia axyridis. Molecular Ecology, 23, 5979-5997. doi: https://doi.org/10.1111/mec.12989 Mack, R.N., Simberloff, D., Lonsdale, M.W., Evans, H., Clout, M., Bazzaz, F.A. (2000) Biotic Invasions : Causes , Epidemiology , Global Consequences , and Control. Ecological Applications, 10, 689-710. doi: https://doi.org/10.1890/1051-0761(2000)010[0689:BICEGC]2.0.CO;2 Mallez, S., Castagnone, C., Lombaert, E., Castagnone-Sereno, P. and Guillemaud, T. (2021) Inference of the worldwide invasion routes of the pinewood nematode Bursaphelenchus xylophilus using approximate Bayesian computation analysis. bioRxiv, 452326, ver. 6 peer-reviewed and recommended by Peer community in Zoology. doi: https://doi.org/10.1101/452326 | Inference of the worldwide invasion routes of the pinewood nematode Bursaphelenchus xylophilus using approximate Bayesian computation analysis | Sophie Mallez, Chantal Castagnone, Eric Lombaert, Philippe Castagnone-Sereno, Thomas Guillemaud | <p>Population genetics have been greatly beneficial to improve knowledge about biological invasions. Model-based genetic inference methods, such as approximate Bayesian computation (ABC), have brought this improvement to a higher level and are now... | | Biogeography, Biological invasions, Ecology, Evolution, Genetics/Genomics, Herbivores, Invertebrates, Molecular biology, Nematology | Stéphane Dupas | 2020-09-15 10:59:41 | ||
25 Aug 2022

Improving species conservation plans under IUCN's One Plan Approach using quantitative genetic methodsQuantitative genetics for a more qualitative conservationRecommended by Peter Galbusera based on reviews by Timothée Bonnet and 1 anonymous reviewerGenetic (bio)diversity is one of three recognised levels of biodiversity, besides species and ecosystem diversity. Its importance for species survival and adaptation is increasingly highlighted and its monitoring recommended (e.g. O’Brien et al 2022). Especially the management of ex-situ populations has a long history of taking into account genetic aspects (through pedigree analysis but increasingly also by applying molecular tools). As in-situ and ex-situ efforts are nowadays often aligned (in a One-Plan-Approach), genetic management is becoming more the standard (supported by quickly developing genomic techniques). However, rarely quantitative genetic aspects are raised in this issue, while its relevance cannot be underestimated. Hence, the current manuscript by Sauve et al (2022) is a welcome contribution, in order to improve conservation efforts. The authors give a clear overview on how quantitative genetic analysis can aid the measurement, monitoring, prediction and management of adaptive genetic variation. The main tools are pedigrees (mainly of ex-situ populations) and the Animal Model. The main goal is to prevent adaption to captivity and altered genetics in general (in reintroduction projects). The confounding factors to take into account (like inbreeding, population structure, differences between facilities, sample size and parental/social effects) are well described by the authors. As such, I fully recommend this manuscript for publication, hoping increased interest in quantitative analysis will benefit the quality of species conservation management. References O'Brien D, Laikre L, Hoban S, Bruford MW et al. (2022) Bringing together approaches to reporting on within species genetic diversity. Journal of Applied Ecology, 00, 1–7. https://doi/10.1111/1365-2664.14225 Sauve D., Spero J., Steiner J., Wheeler H., Lynch C., Chabot A.A. (2022) Improving species conservation plans under IUCN’s One Plan Approach using quantitative genetic methods. EcoEvoRxiv, ver. 9 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.32942/osf.io/n3zxp | Improving species conservation plans under IUCN's One Plan Approach using quantitative genetic methods | Drew Sauve, Jane Hudecki, Jessica Steiner, Hazel Wheeler, Colleen Lynch, Amy A. Chabot | <p>Human activities are resulting in altered environmental conditions that are impacting the demography and evolution of species globally. If we wish to prevent anthropogenic extinction and extirpation, we need to improve our ability to restore wi... | | Conservation biology, Ecology, Evolution, Genetics/Genomics | Peter Galbusera | 2022-02-21 10:45:22 | ||
09 Jul 2021

First detection of herpesvirus and mycoplasma in free-ranging Hermann tortoises (Testudo hermanni), and in potential pet vectorsWelfare threatened speciesRecommended by Peter Galbusera based on reviews by Francis Vercammen and Maria Luisa MarenzoniWildlife is increasingly threatened by drops in number of individuals and populations, and eventually by extinction. Besides loss of habitat, persecution, pet trade,… a decrease in individual health status is an important factor to consider. In this article, Ballouard et al (2021) perform a thorough analysis on the prevalence of two pathogens (herpes virus and mycoplasma) in (mainly) Western Hermann’s tortoises in south-east France. This endangered species was suspected to suffer from infections obtained through released/escaped pet tortoises. By incorporating samples of captive as well as wild tortoises, they convincingly confirm this and identify some possible ‘pet’ vectors. In February this year, a review paper on health assessments in wildlife was published (Kophamel et al 2021). Amongst others, it shows reptilia/chelonia are relatively well-represented among publications. It also contains a useful conceptual framework, in order to improve the quality of the assessments to better facilitate conservation planning. The recommended manuscript (Ballouard et al 2021) adheres to many aspects of this framework (e.g. minimum sample size, risk status, …) while others might need more (future) attention. For example, climate/environmental changes are likely to increase stress levels, which could lead to more disease symptoms. So, follow-up studies should consider conducting endocrinological investigations to estimate/monitor stress levels. Kophamel et al (2021) also stress the importance of strategic international collaboration, which may allow more testing of Eastern Hermann’s Tortoise, as these were shown to be infected by mycoplasma. The genetic health of individuals/populations shouldn’t be forgotten in health/stress assessments. As noted by Ballouard et al (2021), threatened species often have low genetic diversity which makes them more vulnerable to diseases. So, it would be interesting to link the infection data with (individual) genetic characteristics. In future research, the samples collected for this paper could fit that purpose. Finally, it is expected that this paper will contribute to the conservation management strategy of the Hermann’s tortoises. As such, it will be interesting to see how the results of the current paper will be implemented in the ‘field’. As the infections are likely caused by releases/escaped pets and as treating the wild animals is difficult, preventing them from getting infected through pets seems a priority. Awareness building among pet holders and monitoring/treating pets should be highly effective. References Ballouard J-M, Bonnet X, Jourdan J, Martinez-Silvestre A, Gagno S, Fertard B, Caron S (2021) First detection of herpesvirus and mycoplasma in free-ranging Hermann’s tortoises (Testudo hermanni), and in potential pet vectors. bioRxiv, 2021.01.22.427726, ver. 4 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.1101/2021.01.22.427726 Kophamel S, Illing B, Ariel E, Difalco M, Skerratt LF, Hamann M, Ward LC, Méndez D, Munns SL (2021), Importance of health assessments for conservation in noncaptive wildlife. Conservation Biology. https://doi.org/10.1111/cobi.13724 | First detection of herpesvirus and mycoplasma in free-ranging Hermann tortoises (Testudo hermanni), and in potential pet vectors | Jean-marie Ballouard, Xavier Bonnet, Julie Jourdan, Albert Martinez-Silvestre, Stephane Gagno, Brieuc Fertard, Sebastien Caron | <p style="text-align: justify;">Two types of pathogens cause highly contagious upper respiratory tract diseases (URTD) in Chelonians: testudinid herpesviruses (TeHV) and a mycoplasma (<em>Mycoplasma agassizii</em>). In captivity, these infections ... | | Parasitology, Reptiles | Peter Galbusera | 2021-01-25 17:25:34 | ||
05 Jan 2021

Do substrate roughness and gap distance impact gap-bridging strategies in arboreal chameleons?Gap-bridging strategies in arboreal chameleonsRecommended by Ellen Decaestecker based on reviews by Simon Baeckens and 2 anonymous reviewersUntil now, very little is known about the tail use and functional performance in tail prehensile animals. Luger et al. (2020) are the first to provide explorative observations on trait related modulation of tail use, despite the lack of a sufficiently standardized data set to allow statistical testing. They described whether gap distance, perch diameter, and perch roughness influence tail use and overall locomotor behavior of the species Chamaeleo calyptratus. References Luger, A.M., Vermeylen, V., Herrel, A. and Adriaens, D. (2020) Do substrate roughness and gap distance impact gap-bridging strategies in arboreal chameleons? bioRxiv, 2020.08.21.260596, ver. 3 peer-reviewed and recommended by PCI Zoology. doi: https://doi.org/10.1101/2020.08.21.260596 | Do substrate roughness and gap distance impact gap-bridging strategies in arboreal chameleons? | Allison M. Luger, Vincent Vermeylen, Anthony Herrel, Dominique Adriaens | <p>Chameleons are well-equipped for an arboreal lifestyle, having ‘zygodactylous’ hands and feet as well as a fully prehensile tail. However, to what degree tail use is preferred over autopod prehension has been largely neglected. Using an indoor ... | | Behavior, Biology, Herpetology, Reptiles, Vertebrates | Ellen Decaestecker | 2020-08-25 10:06:42 | ||
24 Jun 2022

Dopamine pathway characterization during the reproductive mode switch in the pea aphidIn search of the links between environmental signals and polyphenismRecommended by Mathieu Joron based on reviews by Antonia Monteiro and 2 anonymous reviewersPolyphenisms offer an opportunity to study the links between phenotype, development, and environment in a controlled genomic context (Simpson, Sword, & Lo, 2011). In organisms with short generation times, individuals living and developing in different seasons encounter different environmental conditions. Adaptive plasticity allows them to express different phenotypes in response to seasonal cues, such as temperature or photoperiod. Such phenotypes can be morphological variants, for instance displaying different wing patterns as seen in butterflies (Brakefield & Larsen, 1984; Nijhout, 1991; Windig, 1999), or physiological variants, characterized for instance by direct development vs winter diapause in temperate insects (Dalin & Nylin, 2012; Lindestad, Wheat, Nylin, & Gotthard, 2019; Shearer et al., 2016). Many aphids display cyclical parthenogenesis, a remarkable seasonal polyphenism for reproductive mode (Tagu, Sabater-Muñoz, & Simon, 2005), also sometimes coupled with wing polyphenism (Braendle, Friebe, Caillaud, & Stern, 2005), which allows them to switch between parthenogenesis during spring and summer to sexual reproduction and the production of diapausing eggs before winter. In the pea aphid Acyrthosiphon pisum, photoperiod shortening results in the production, by parthenogenetic females, of embryos developing into the parthenogenetic mothers of sexual individuals. The link between parthenogenetic reproduction and sexual reproduction, therefore, occurs over two generations, changing from a parthenogenetic form producing parthenogenetic females (virginoparae), to a parthenogenetic form producing sexual offspring (sexuparae), and finally sexual forms producing overwintering eggs (Le Trionnaire et al., 2022). The molecular basis for the transduction of the environmental signal into reproductive changes is still unknown, but the dopamine pathway is an interesting candidate. Form-specific expression of certain genes in the dopamine pathway occurs downstream of the perception of the seasonal cue, notably with a marked decrease in the heads of embryos reared under short-day conditions and destined to become sexuparae. Dopamine has multiple roles during development, with one mode of action in cuticle melanization and sclerotization, and a neurological role as a synaptic neurotransmitter. Both modes of action might be envisioned to contribute functionally to the perception and transduction of environmental signals. In this study, Le Trionnaire and colleagues aim at clarifying this role in the pea aphid (Le Trionnaire et al., 2022). Using quantitative RT-PCR, RNA-seq, and in situ hybridization of RNA probes, they surveyed the timing and spatial patterns of expression of dopamine pathway genes during the development of different stages of embryo to larvae reared under long and short-day conditions, and destined to become virginoparae or sexuparae females, respectively. The genes involved in the synaptic release of dopamine generally did not show differences in expression between photoperiodic treatments. By contrast, pale and ddc, two genes acting upstream of dopamine production, generally tended to show a downregulation in sexuparare embryo, as well as genes involved in cuticle development and interacting with the dopamine pathway. The downregulation of dopamine pathway genes observed in the heads of sexuparare juveniles is already detectable at the embryonic stage, suggesting embryos might be sensing environmental cues leading them to differentiate into sexuparae females. The way pale and ddc expression differences could influence environmental sensitivity is still unclear. The results suggest that a cuticle phenotype specifically in the heads of larvae could be explored, perhaps in the form of a reduction in cuticle sclerotization and melanization which might allow photoperiod shortening to be perceived and act on development. Although its causality might be either way, such a link would be exciting to investigate, yet the existence of cuticle differences between the two reproductive types is still a hypothesis to be tested. The lack of differences in the expression of synaptic release genes for dopamine might seem to indicate that the plastic response to photoperiod is not mediated via neurological roles. Yet, this does not rule out the role of decreasing levels of dopamine in mediating this response in the central nervous system of embryos, even if the genes regulating synaptic release are equally expressed. To test for a direct role of ddc in regulating the reproductive fate of embryos, the authors have generated CrispR-Cas9 knockout mutants. Those mutants displayed egg cuticle melanization, but with lethal effects, precluding testing the effect of ddc at later stages in development. Gene manipulation becomes feasible in the pea aphid, opening up certain avenues for understanding the roles of other genes during development. This study adds nicely to our understanding of the intricate changes in gene expression involved in polyphenism. But it also shows the complexity of deciphering the links between environmental perception and changes in physiology, which mobilise multiple interacting gene networks. In the era of manipulative genetics, this study also stresses the importance of understanding the traits and phenotypes affected by individual genes, which now seems essential to piece the puzzle together. References Braendle C, Friebe I, Caillaud MC, Stern DL (2005) Genetic variation for an aphid wing polyphenism is genetically linked to a naturally occurring wing polymorphism. Proceedings of the Royal Society B: Biological Sciences, 272, 657–664. https://doi.org/10.1098/rspb.2004.2995 Brakefield PM, Larsen TB (1984) The evolutionary significance of dry and wet season forms in some tropical butterflies. Biological Journal of the Linnean Society, 22, 1–12. https://doi.org/10.1111/j.1095-8312.1984.tb00795.x Dalin P, Nylin S (2012) Host-plant quality adaptively affects the diapause threshold: evidence from leaf beetles in willow plantations. Ecological Entomology, 37, 490–499. https://doi.org/10.1111/j.1365-2311.2012.01387.x Le Trionnaire G, Hudaverdian S, Richard G, Tanguy S, Gleonnec F, Prunier-Leterme N, Gauthier J-P, Tagu D (2022) Dopamine pathway characterization during the reproductive mode switch in the pea aphid. bioRxiv, 2020.03.10.984989, ver. 4 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.1101/2020.03.10.984989 Lindestad O, Wheat CW, Nylin S, Gotthard K (2019) Local adaptation of photoperiodic plasticity maintains life cycle variation within latitudes in a butterfly. Ecology, 100, e02550. https://doi.org/10.1002/ecy.2550 Nijhout HF (1991). The development and evolution of butterfly wing patterns. Washington, DC: Smithsonian Institution Press. Shearer PW, West JD, Walton VM, Brown PH, Svetec N, Chiu JC (2016) Seasonal cues induce phenotypic plasticity of Drosophila suzukii to enhance winter survival. BMC Ecology, 16, 11. https://doi.org/10.1186/s12898-016-0070-3 Simpson SJ, Sword GA, Lo N (2011) Polyphenism in Insects. Current Biology, 21, R738–R749. https://doi.org/10.1016/j.cub.2011.06.006 Tagu D, Sabater-Muñoz B, Simon J-C (2005) Deciphering reproductive polyphenism in aphids. Invertebrate Reproduction & Development, 48, 71–80. https://doi.org/10.1080/07924259.2005.9652172 Windig JJ (1999) Trade-offs between melanization, development time and adult size in Inachis io and Araschnia levana (Lepidoptera: Nymphalidae)? Heredity, 82, 57–68. https://doi.org/10.1038/sj.hdy.6884510 | Dopamine pathway characterization during the reproductive mode switch in the pea aphid | Gaël Le Trionnaire, Sylvie Hudaverdian, Gautier Richard, Sylvie Tanguy, Florence Gleonnec, Nathalie Prunier-Leterme, Jean-Pierre Gauthier, Denis Tagu | <p>Aphids are major pests of most of the crops worldwide. Such a success is largely explained by the remarkable plasticity of their reproductive mode. They reproduce efficiently by viviparous parthenogenesis during spring and summer generating imp... | | Development, Genetics/Genomics, Insecta, Molecular biology | Mathieu Joron | 2020-03-13 13:01:44 | ||
10 Jan 2020

Culex saltanensis and Culex interfor (Diptera: Culicidae) are susceptible and competent to transmit St. Louis encephalitis virus (Flavivirus: Flaviviridae) in central ArgentinaMultiple vector species may be responsible for transmission of Saint Louis Encephalatis Virus in ArgentinaRecommended by Anna Cohuet based on reviews by 2 anonymous reviewersMedical and veterinary entomology is a discipline that deals with the role of insects on human and animal health. A primary objective is the identification of vectors that transmit pathogens. This is the aim of Beranek and co-authors in their study [1]. They focus on mosquito vector species responsible for transmission of St. Louis encephalitis virus (SLEV), an arbovirus that circulates in avian species but can incidentally occur in dead end mammal hosts such as humans, inducing symptoms and sometimes fatalities. Culex pipiens quinquefasciatus is known as the most common vector, but other species are suspected to also participate in transmission. Among them Culex saltanensis and Culex interfor have been found to be infected by the virus in the context of outbreaks. The fact that field collected mosquitoes carry virus particles is not evidence for their vector competence: indeed to be a competent vector, the mosquito must not only carry the virus, but also the virus must be able to replicate within the vector, overcome multiple barriers (until the salivary glands) and be present at sufficient titre within the saliva. This paper describes the experiments implemented to evaluate the vector competence of Cx. saltanensis and Cx. interfor from ingestion of SLEV to release within the saliva. Females emerged from field-collected eggs of Cx. pipiens quinquefasciatus, Cx. saltanensis and Cx. interfor were allowed to feed on SLEV infected chicks and viral development was measured by using (i) the infection rate (presence/absence of virus in the mosquito abdomen), (ii) the dissemination rate (presence/absence of virus in mosquito legs), and (iii) the transmission rate (presence/absence of virus in mosquito saliva). The sample size for each species is limited because of difficulties for collecting, feeding and maintaining large numbers of individuals from field populations, however the results are sufficient to show that this strain of SLEV is able to disseminate and be expelled in the saliva of mosquitoes of the three species at similar viral loads. This work therefore provides evidence that Cx saltanensis and Cx interfor are competent species for SLEV to complete its life-cycle. Vector competence does not directly correlate with the ability to transmit in real life as the actual vectorial capacity also depends on the contact between the infectious vertebrate hosts, the mosquito life expectancy and the extrinsic incubation period of the viruses. The present study does not deal with these characteristics, which remain to be investigated to complete the picture of the role of Cx saltanensis and Cx interfor in SLEV transmission. However, this study provides proof of principle that that SLEV can complete it’s life-cycle in Cx saltanensis and Cx interfor. Combined with previous knowledge on their feeding preference, this highlights their potential role as bridge vectors between birds and mammals. These results have important implications for epidemiological forecasting and disease management. Public health strategies should consider the diversity of vectors in surveillance and control of SLEV. References | Culex saltanensis and Culex interfor (Diptera: Culicidae) are susceptible and competent to transmit St. Louis encephalitis virus (Flavivirus: Flaviviridae) in central Argentina | Beranek MD, Quaglia AI, Peralta GC, Flores FS, Stein M, Diaz LA, Almirón WR and Contigiani MS | <p>Infectious diseases caused by mosquito-borne viruses constitute health and economic problems worldwide. St. Louis encephalitis virus (SLEV) is endemic and autochthonous in the American continent. Culex pipiens quinquefasciatus is the primary ur... | | Medical entomology | Anna Cohuet | 2019-08-03 00:56:38 | ||
02 Nov 2021

Cuckoo male bumblebees perform slower and longer flower visits than free-living male and worker bumblebeesCuckoo bumblebee males might reduce plant fitnessRecommended by Michael Lattorff based on reviews by Patrick Lhomme, Silvio Erler and 2 anonymous reviewers
In pollinator insects, especially bees, foraging is almost exclusively performed by females due to the close linkage with brood care. They collect pollen as a protein- and lipid-rich food to feed developing larvae in solitary and social species. Bees take carbohydrate-rich nectar in small quantities to fuel their flight and carry the pollen load. To optimise the foraging flight, they tend to be flower constant, reducing the flower handling time and time among individual inflorescences (Goulson, 1999). Males of pollinator species might be found on flowers as well. As they do not collect any pollen for brood care, their foraging flights and visits to flowers might not be shaped by the selective forces that optimise the foraging flights of females. They might stay longer in individual flowers, take up nectar if needed, but might unintentionally carry pollen on their body surface (Wolf & Moritz, 2014). | Cuckoo male bumblebees perform slower and longer flower visits than free-living male and worker bumblebees | Alessandro Fisogni, Gherardo Bogo, François Massol, Laura Bortolotti, Marta Galloni | <p>Cuckoo bumblebees are a monophyletic group within the genus Bombus and social parasites of free-living bumblebees, upon which they rely to rear their offspring. Cuckoo bumblebees lack the worker caste and visit flowers primarily for their own s... | | Behavior, Biology, Ecology, Insecta, Invertebrates, Terrestrial | Michael Lattorff | Patrick Lhomme, Seth Barribeau , Silvio Erler, Denis Michez | 2021-02-02 01:41:35 |




