
Latest recommendations

| Id | Title * | Authors * | Abstract * | Picture * | Thematic fields * | Recommender▼ | Reviewers | Submission date | |
|---|---|---|---|---|---|---|---|---|---|
21 Mar 2023
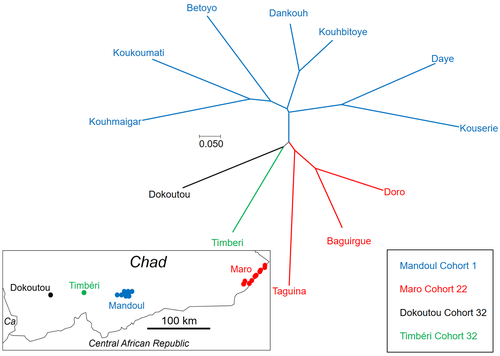
Population genetics of Glossina fuscipes fuscipes from southern ChadPopulation genetics of tsetse, the vector of African Trypanosomiasis, helps informing strategies for control programsRecommended by Michael Lattorff based on reviews by 2 anonymous reviewers based on reviews by 2 anonymous reviewers
Human African Trypanosomiasis (HAT), or sleeping sickness, is caused by trypanosome parasites. In sub-Saharan Africa, two forms are present, Trypanosoma brucei gambiense and T. b. rhodesiense, the former responsible for 95% of reported cases. The parasites are transmitted through a vector, Genus Glossina, also known as tsetse, which means fly in Tswana, a language from southern Africa. Through a blood meal, tsetse picks up the parasite from infected humans or animals (in animals, the parasite causes Animal African Trypanosomiasis or nagana disease). Through medical interventions and vector control programs, the burden of the disease has drastically reduced over the past two decades, so the WHO neglected tropical diseases road map targets the interruption of transmission (zero cases) for 2030 (WHO 2022). Meaningful vector control programs utilize traps for the removal of animals and for surveillance, along with different methods of spraying insecticides. However, in existing HAT risk areas, it will be essential to understand the ecology of the vector species to implement control programs in a way that areas cleared from the vector will not be reinvaded from other populations. Thus, it will be crucial to understand basic population genetics parameters related to population structure and subdivision, migration frequency and distances, population sizes, and the potential for sex-biased dispersal. The authors utilize genotyping using nine highly polymorphic microsatellite markers of samples from Chad collected in differently affected regions and at different time points (Ravel et al., 2023). Two major HAT zones exist that are targeted by vector control programs, namely Madoul and Maro, while two other areas, Timbéri and Dokoutou, are free of trypanosomes. Samples were taken before vector control programs started. The sex ratio was female-biased, most strongly in Mandoul and Maro, the zones with the lowest population density. This could be explained by resource limitation, which could be the hosts for a blood meal or the sites for larviposition. Limited resources mean that females must fly further, increasing the chance that more females are caught in traps. The effective population densities of Mandoul and Maro were low. However, there was a convergence of population density and trapping density, which might be explained by the higher preservation of flies in the high-density areas of Timbéri and Dokoutou after the first round of sampling, which can only be tested using a second sampling. The dispersal distances are the highest recorded so far, especially in Mandoul and Maro, with 20-30 km per generation. However, in Timbéri and Dokoutou, which are 50 km apart, very little exchange occurs (approx. 1-2 individuals every six months). A major contributor to this is the massive destruction of habitat that started in the early 1990s and left patchily distributed and fragmented habitats. The Mandoul zone might be safe from reinvasion after eradication, as for a successful re-establishment, either a pair of a female and male or a pregnant female are required. As the trypanosome prevalence amongst humans was 0.02 and of tsetse 0.06 (Ibrahim et al., 2021) before interventions began, medical interventions and vector control might have further reduced these levels, making a reinvasion and subsequent re-establishment of HAT very unlikely. Maro is close to the border of the Central African Republic, and the area has not been well investigated concerning refugee populations of tsetse, which could contribute to a reinvasion of the Maro zone. The higher level of genetic heterogeneity of the Maro population indicates that invasions from neighboring populations are already ongoing. This immigration could also be the reason for not detecting the bottleneck signature in the Maro population. The two HAT areas need different levels of attention while implementing vector eradication programs. While Madoul is relatively safe against reinvasion, Maro needs another type of attention, as frequent and persistent immigration might counteract eradication efforts. Thus, it is recommended that continuous tsetse suppression needs to be implemented in Maro. This study shows nicely that an in-depth knowledge of the processes within and between populations is needed to understand how these populations behave. This can be used to extrapolate, make predictions, and inform the organisations implementing vector control programs to include valuable adjustments, as in the case of Maro. Such integrative approaches can help prevent the failure of programs, potentially saving costs and preventing infections of humans and animals who might die if not treated. References Ibrahim MAM, Weber JS, Ngomtcho SCH, Signaboubo D, Berger P, Hassane HM, Kelm S (2021) Diversity of trypanosomes in humans and cattle in the HAT foci Mandoul and Maro, Southern Chad- Southern Chad-A matter of concern for zoonotic potential? PLoS Neglected Tropical Diseases, 15, e000 323. https://doi.org/10.1371/journal.pntd.0009323 Ravel S, Mahamat MH, Ségard A, Argiles-Herrero R, Bouyer J, Rayaisse JB, Solano P, Mollo BG, Pèka M, Darnas J, Belem AMG, Yoni W, Noûs C, de Meeûs T (2023) Population genetics of Glossina fuscipes fuscipes from southern Chad. Zenodo, ver. 9 peer-reviewed and recommended by PCI Zoology. https://doi.org/10.5281/zenodo.7763870 WHO (2022) Trypanosomiasis, human African (sleeping sickness). https://www.who.int/news-room/fact-sheets/detail/trypanosomiasis-human-african-(sleeping-sickness), retrieved 17. March 2023 | Population genetics of Glossina fuscipes fuscipes from southern Chad | Sophie Ravel, Mahamat Hissène Mahamat, Adeline Ségard, Rafael Argiles-Herrero, Jérémy Bouyer, Jean-Baptiste Rayaisse, Philippe Solano, Brahim Guihini Mollo, Mallaye Pèka, Justin Darnas, Adrien Marie Gaston Belem, Wilfrid Yoni, Camille Noûs, Thierr... | <p>In Subsaharan Africa, tsetse flies (genus Glossina) are vectors of trypanosomes causing Human African Trypanosomiasis (HAT) and Animal African Trypanosomosis (AAT). Some foci of HAT persist in Southern Chad, where a program of tsetse control wa... | | Biology, Ecology, Evolution, Genetics/Genomics, Insecta, Medical entomology, Parasitology, Pest management, Veterinary entomology | Michael Lattorff | Audrey Bras | 2022-04-22 11:25:24 | |
27 Apr 2023
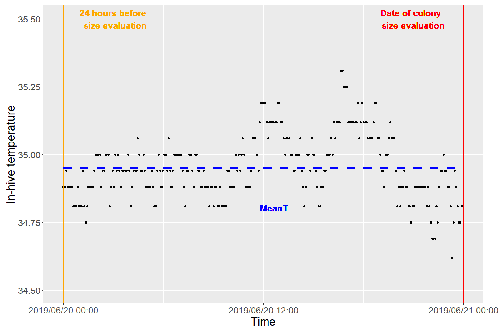
Brood thermoregulation effectivenessis positively linked to the amount of brood but not to the number of bees in honeybee coloniesPrecision and accuracy of honeybee thermoregulationRecommended by Michael Lattorff based on reviews by Jakob Wegener and Christopher Mayack
The Western honeybee, Apis mellifera L., is one of the best-studied social insects. It shows a reproductive division of labour, cooperative brood care, and age-related polyethism. Furthermore, honeybees regulate the temperature in the hive. Although bees are invertebrates that are usually ectothermic, this is still true for individual worker bees, but the colony maintains a very narrow range of temperature, especially within the brood nest. This is quite important as the development of individuals is dependent on ambient temperature, with higher temperatures resulting in accelerated development and vice versa. In honeybees, a feedback mechanism couples developmental temperature and the foraging behaviour of the colony and the future population development (Tautz et al., 2003). Bees raised under lower temperatures are more likely to perform in-hive tasks, while bees raised under higher temperatures are better foragers. To maintain optimal levels of worker population growth and foraging rates, it is adaptive to regulate temperature to ensure optimal levels of developing brood. Moreover, this allows honeybees to decouple the internal developmental processes from ambient temperatures enhancing the ecological success of the species. In every system of thermoregulation, whether it is endothermic under the utilization of energetic resources as in mammals or the honeybee or ectothermic as in lower vertebrates and invertebrates through differential exposure to varying environmental temperature gradients, there is a need for precision (low variability) and accuracy (hitting the target temperature). However, in honeybees, the temperature is regulated by workers through muscle contraction and fanning of the wings and thus, a higher number of workers could be better at achieving precise and accurate temperature within the brood nest. Alternatively, the amount of brood could trigger responses with more brood available, a need for more precise and accurate temperature control. The authors aimed at testing these two important factors on the precision and accuracy of within-colony temperature regulation by monitoring 28 colonies equipped with temperature sensors for two years (Godeau et al., 2023). They found that the number of brood cells predicted the mean temperature (accuracy of thermoregulation). Other environmental factors had a small effect. However, the model incorporating these factors was weak in predicting the temperature as it overestimated temperatures in lower ranges and underestimated temperatures in higher ranges. In contrast, the variability of the target temperature (precision of thermoregulation) was positively affected by the external temperature, while all other factors did not show a significant effect. Again, the model was weak in predicting the data. Overall colony size measured in categories of the number of workers and the number of brood cells did not show major differences in variability of the mean temperature, but a slight positive effect for the number of bees on the mean temperature. Unfortunately, the temperature was a poor predictor of colony size. The latter is important as the remote control of beehives using Internet of Things (IoT) technologies get more and more incorporated into beekeeping management. These IoT technologies and their success are dependent on good proxies for the control of the status of the colony. Amongst the factors to monitor, the colony size (number of bees and/or amount of brood) is extremely important, but temperature measurements alone will not allow us to predict colony sizes. Nevertheless, this study showed clearly that the number of brood cells is a crucial factor for the accuracy of thermoregulation in the beehive, while ambient temperature affects the precision of thermoregulation. In the view of climate change, the latter factor seems to be important, as more extreme environmental conditions in the future call for measures of mitigation to ensure the proper functioning of the bee colony, including the maintenance of homeostatic conditions inside of the nest to ensure the delivery of the ecosystem service of pollination. REFERENCES Godeau U, Pioz M, Martin O, Rüger C, Crauser D, Le Conte Y, Henry M, Alaux C (2023) Brood thermoregulation effectiveness is positively linked to the amount of brood but not to the number of bees in honeybee colonies. EcoEvoRxiv, ver. 5 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.32942/osf.io/9mwye Tautz J, Maier S, Claudia Groh C, Wolfgang Rössler W, Brockmann A (2003) Behavioral performance in adult honey bees is influenced by the temperature experienced during their pupal development. PNAS 100: 7343–7347. https://doi.org/10.1073/pnas.1232346100 | Brood thermoregulation effectivenessis positively linked to the amount of brood but not to the number of bees in honeybee colonies | Ugoline Godeau, Maryline Pioz, Olivier Martin, Charlotte Rüger, Didier Crauser, Yves Le Conte, Mickael Henry, Cédric Alaux | <p style="text-align: justify;">To ensure the optimal development of brood, a honeybee colony needs to regulate its temperature within a certain range of values (thermoregulation), regardless of environmental changes in biotic and abiotic factors.... | | Biology, Conservation biology, Demography/population dynamics, Ecology, Insecta | Michael Lattorff | Mauricio Daniel Beranek | 2022-07-06 09:20:10 | |
30 Nov 2022
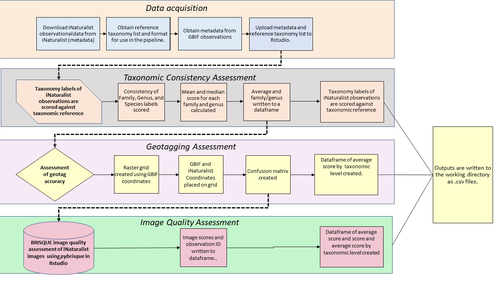
A pipeline for assessing the quality of images and metadata from crowd-sourced databases.Harnessing the full potential of iNaturalist and other databasesRecommended by Matthias Foellmer based on reviews by Clive Hambler and Catherine ScottThe popularity of iNaturalist and other online biodiversity databases to which the general public and specialists alike contribute observations has skyrocketed in recent years (Dance 2022). The AI-based algorithms (computer vision) which provide the first identification of a given organism on an uploaded photograph have become very sophisticated, suggesting initial identifications often down to species level with a surprisingly high degree of accuracy. The initial identifications are then confirmed or improved by feedback from the community, which works particularly well for organismal groups to which many active community members contribute, such as the birds. Hence, providing initial observations and identifying observations of others, as well as browsing the recorded biodiversity for given locales or the range of occurrences of individual taxa has become a meaningful and satisfying experience for the interested naturalist. Furthermore, several research studies have now been published relying on observations uploaded to iNaturalist (Szentivanyi and Vincze 2022). However, using the enormous amount of natural history data available on iNaturalist in a systematic way has remained challenging, since this requires not only retrieving numerous observations from the database (in the hundreds or even thousands), but also some level of transparent quality control. Billotte (2022) provides a protocol and R scripts for the quality assessment of downloaded observations from iNaturalist, allowing an efficient and reproducible stepwise approach to prepare a high-quality data set for further analysis. First, observations with their associated metadata are downloaded from iNaturalist, along with the corresponding entries from the Global Biodiversity Information Facility (GBIF). In addition, a taxonomic reference list is obtained (these are available online for many taxa), which is used to assess the taxonomic consistency in the dataset. Second, the geo-tagging is assessed by comparing the iNaturalist and GBIF metadata. Lastly, the image quality is assessed using pyBRISQUE. The approach is illustrated using spiders (Araneae) as an example. Spiders are a very diverse taxon and an excellent taxonomic reference list is available (World Spider Catalogue 2022). However, spiders are not well known to most non-specialists, and it is not easy to take good pictures of spiders without using professional equipment. Therefore, the ability of iNaturalist’s computer vision to provide identifications is limited to this date and the community of specialists active on iNaturalist is comparatively small. Hence, spiders are a good taxon to demonstrate how the pipeline results in a quality-controlled dataset based on crowed-sourced data. Importantly, the software employed is free to use, although inevitably, the initial learning curve to use R scripts can be steep, depending on prior expertise with R/RStudio. Furthermore, the approach is employable with databases other than iNaturalist. In summary, Billotte's (2022) pipeline allows researchers to use the wealth of observations on iNaturalist and other databases to produce large metadata and image datasets of high-quality in a reproducible way. This should pave the way for more studies, which could include, for example, the assessment of range expansions of invasive species or the evaluation of the presence of endangered species, potentially supporting conservation efforts. References Billotte J (2022) A pipeline for assessing the quality of images and metadata from crowd-sourced databases. BiorXiv, 2022.04.29.490112, ver 5 peer reviewed and recommended by Peer Community In Zoology. https://doi.org/10.1101/2022.04.29.490112 Dance A (2022) Community science draws on the power of the crowd. Nature, 609, 641–643. https://doi.org/10.1038/d41586-022-02921-3 Szentivanyi T, Vincze O (2022) Tracking wildlife diseases using community science: an example through toad myiasis. European Journal of Wildlife Research, 68, 74. https://doi.org/10.1007/s10344-022-01623-5 World Spider Catalog (2022). World Spider Catalog. Version 23.5. Natural History Museum Bern, online at http://wsc.nmbe.ch. https://doi.org/10.24436/2 | A pipeline for assessing the quality of images and metadata from crowd-sourced databases. | Jackie Billotte | <p style="text-align: justify;">Crowd-sourced biodiversity databases provide easy access to data and images for ecological education and research. One concern with using publicly sourced databases; however, is the quality of their images, taxonomi... | | Arachnids, Biodiversity, Biology, Conservation biology, Ecology, Insecta, Invertebrates | Matthias Foellmer | 2022-05-03 00:18:23 | ||
24 Jun 2022

Dopamine pathway characterization during the reproductive mode switch in the pea aphidIn search of the links between environmental signals and polyphenismRecommended by Mathieu Joron based on reviews by Antonia Monteiro and 2 anonymous reviewersPolyphenisms offer an opportunity to study the links between phenotype, development, and environment in a controlled genomic context (Simpson, Sword, & Lo, 2011). In organisms with short generation times, individuals living and developing in different seasons encounter different environmental conditions. Adaptive plasticity allows them to express different phenotypes in response to seasonal cues, such as temperature or photoperiod. Such phenotypes can be morphological variants, for instance displaying different wing patterns as seen in butterflies (Brakefield & Larsen, 1984; Nijhout, 1991; Windig, 1999), or physiological variants, characterized for instance by direct development vs winter diapause in temperate insects (Dalin & Nylin, 2012; Lindestad, Wheat, Nylin, & Gotthard, 2019; Shearer et al., 2016). Many aphids display cyclical parthenogenesis, a remarkable seasonal polyphenism for reproductive mode (Tagu, Sabater-Muñoz, & Simon, 2005), also sometimes coupled with wing polyphenism (Braendle, Friebe, Caillaud, & Stern, 2005), which allows them to switch between parthenogenesis during spring and summer to sexual reproduction and the production of diapausing eggs before winter. In the pea aphid Acyrthosiphon pisum, photoperiod shortening results in the production, by parthenogenetic females, of embryos developing into the parthenogenetic mothers of sexual individuals. The link between parthenogenetic reproduction and sexual reproduction, therefore, occurs over two generations, changing from a parthenogenetic form producing parthenogenetic females (virginoparae), to a parthenogenetic form producing sexual offspring (sexuparae), and finally sexual forms producing overwintering eggs (Le Trionnaire et al., 2022). The molecular basis for the transduction of the environmental signal into reproductive changes is still unknown, but the dopamine pathway is an interesting candidate. Form-specific expression of certain genes in the dopamine pathway occurs downstream of the perception of the seasonal cue, notably with a marked decrease in the heads of embryos reared under short-day conditions and destined to become sexuparae. Dopamine has multiple roles during development, with one mode of action in cuticle melanization and sclerotization, and a neurological role as a synaptic neurotransmitter. Both modes of action might be envisioned to contribute functionally to the perception and transduction of environmental signals. In this study, Le Trionnaire and colleagues aim at clarifying this role in the pea aphid (Le Trionnaire et al., 2022). Using quantitative RT-PCR, RNA-seq, and in situ hybridization of RNA probes, they surveyed the timing and spatial patterns of expression of dopamine pathway genes during the development of different stages of embryo to larvae reared under long and short-day conditions, and destined to become virginoparae or sexuparae females, respectively. The genes involved in the synaptic release of dopamine generally did not show differences in expression between photoperiodic treatments. By contrast, pale and ddc, two genes acting upstream of dopamine production, generally tended to show a downregulation in sexuparare embryo, as well as genes involved in cuticle development and interacting with the dopamine pathway. The downregulation of dopamine pathway genes observed in the heads of sexuparare juveniles is already detectable at the embryonic stage, suggesting embryos might be sensing environmental cues leading them to differentiate into sexuparae females. The way pale and ddc expression differences could influence environmental sensitivity is still unclear. The results suggest that a cuticle phenotype specifically in the heads of larvae could be explored, perhaps in the form of a reduction in cuticle sclerotization and melanization which might allow photoperiod shortening to be perceived and act on development. Although its causality might be either way, such a link would be exciting to investigate, yet the existence of cuticle differences between the two reproductive types is still a hypothesis to be tested. The lack of differences in the expression of synaptic release genes for dopamine might seem to indicate that the plastic response to photoperiod is not mediated via neurological roles. Yet, this does not rule out the role of decreasing levels of dopamine in mediating this response in the central nervous system of embryos, even if the genes regulating synaptic release are equally expressed. To test for a direct role of ddc in regulating the reproductive fate of embryos, the authors have generated CrispR-Cas9 knockout mutants. Those mutants displayed egg cuticle melanization, but with lethal effects, precluding testing the effect of ddc at later stages in development. Gene manipulation becomes feasible in the pea aphid, opening up certain avenues for understanding the roles of other genes during development. This study adds nicely to our understanding of the intricate changes in gene expression involved in polyphenism. But it also shows the complexity of deciphering the links between environmental perception and changes in physiology, which mobilise multiple interacting gene networks. In the era of manipulative genetics, this study also stresses the importance of understanding the traits and phenotypes affected by individual genes, which now seems essential to piece the puzzle together. References Braendle C, Friebe I, Caillaud MC, Stern DL (2005) Genetic variation for an aphid wing polyphenism is genetically linked to a naturally occurring wing polymorphism. Proceedings of the Royal Society B: Biological Sciences, 272, 657–664. https://doi.org/10.1098/rspb.2004.2995 Brakefield PM, Larsen TB (1984) The evolutionary significance of dry and wet season forms in some tropical butterflies. Biological Journal of the Linnean Society, 22, 1–12. https://doi.org/10.1111/j.1095-8312.1984.tb00795.x Dalin P, Nylin S (2012) Host-plant quality adaptively affects the diapause threshold: evidence from leaf beetles in willow plantations. Ecological Entomology, 37, 490–499. https://doi.org/10.1111/j.1365-2311.2012.01387.x Le Trionnaire G, Hudaverdian S, Richard G, Tanguy S, Gleonnec F, Prunier-Leterme N, Gauthier J-P, Tagu D (2022) Dopamine pathway characterization during the reproductive mode switch in the pea aphid. bioRxiv, 2020.03.10.984989, ver. 4 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.1101/2020.03.10.984989 Lindestad O, Wheat CW, Nylin S, Gotthard K (2019) Local adaptation of photoperiodic plasticity maintains life cycle variation within latitudes in a butterfly. Ecology, 100, e02550. https://doi.org/10.1002/ecy.2550 Nijhout HF (1991). The development and evolution of butterfly wing patterns. Washington, DC: Smithsonian Institution Press. Shearer PW, West JD, Walton VM, Brown PH, Svetec N, Chiu JC (2016) Seasonal cues induce phenotypic plasticity of Drosophila suzukii to enhance winter survival. BMC Ecology, 16, 11. https://doi.org/10.1186/s12898-016-0070-3 Simpson SJ, Sword GA, Lo N (2011) Polyphenism in Insects. Current Biology, 21, R738–R749. https://doi.org/10.1016/j.cub.2011.06.006 Tagu D, Sabater-Muñoz B, Simon J-C (2005) Deciphering reproductive polyphenism in aphids. Invertebrate Reproduction & Development, 48, 71–80. https://doi.org/10.1080/07924259.2005.9652172 Windig JJ (1999) Trade-offs between melanization, development time and adult size in Inachis io and Araschnia levana (Lepidoptera: Nymphalidae)? Heredity, 82, 57–68. https://doi.org/10.1038/sj.hdy.6884510 | Dopamine pathway characterization during the reproductive mode switch in the pea aphid | Gaël Le Trionnaire, Sylvie Hudaverdian, Gautier Richard, Sylvie Tanguy, Florence Gleonnec, Nathalie Prunier-Leterme, Jean-Pierre Gauthier, Denis Tagu | <p>Aphids are major pests of most of the crops worldwide. Such a success is largely explained by the remarkable plasticity of their reproductive mode. They reproduce efficiently by viviparous parthenogenesis during spring and summer generating imp... | | Development, Genetics/Genomics, Insecta, Molecular biology | Mathieu Joron | 2020-03-13 13:01:44 | ||
14 Dec 2023
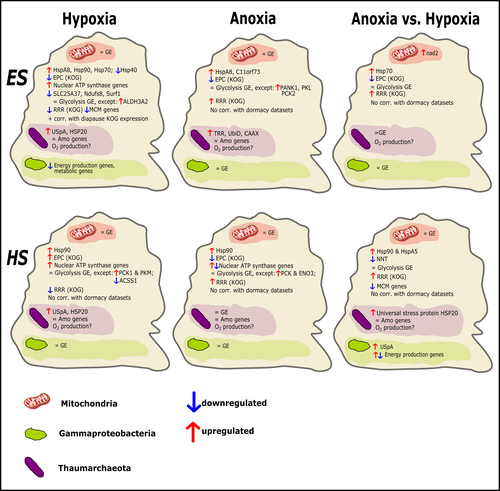
Transcriptomic responses of sponge holobionts to in situ, seasonal anoxia and hypoxiaFuture oceanic conditions could leave sponge holobionts breathless – but they won’t let that stop themRecommended by Loïc N. Michel based on reviews by Maria Lopez Acosta and 2 anonymous reviewers
It is now widely accepted that anthropogenic climate change is a severe threat to biodiversity, ecosystem function and associated ecosystem services. Assessing the vulnerability of species and predicting their response to future changes has become a priority for environmental biology (Williams et al. 2020). Over the last few decades, oxygen concentrations in both the open ocean and coastal waters have been declining steadily as the result of multiple anthropogenic activities. This global trends towards hypoxia is expected to continue in the future, causing a host of negative effects on marine ecosystems. Oxygen is indeed crucial to many biological processes in the ocean, and its decrease could have strong impacts on biogeochemical cycles, and therefore on marine productivity and biodiversity (Breitburg et al. 2018). Whenever facing such drastic environmental changes, all organisms are expected to have some intrinsic ability to adapt. At shorter than evolutionary timescales, ecological plasticity and the eco-physiological processes that sustain it could constitute important adaptive mechanisms (Williams et al. 2020) Marine sponges seem particularly well-adapted to oxygen deficiency, as some species can survive seasonal anoxia for several months. This paper by Strehlow et al. (2023) examines the mechanisms allowing this exceptional tolerance. Focusing on two species of sponges, they used transcriptomics to assess how gene expression by sponges, by their mitochondria, or by their unique and species-specific microbiome could facilitate this trait. Their results suggest that sponge holobionts maintain metabolic activity under anoxic conditions while displaying shock response, therefore not supporting the hypothesis of sponge dormancy. Furthermore, hypoxia and anoxia seemed to influence gene expression in different ways, highlighting the complexity of sponge response to deoxygenation. As often, their exciting results raise as many questions as they provide answers and pave the way for more research regarding how anoxia tolerance in marine sponges could give them an advantage in future oceanic environmental conditions. References Breitburg et al. (2018): Declining oxygen in the global ocean and coastal waters. Science 359, eaam7240. https://doi.org/10.1126/science.aam7240 Strehlow et al. (2023): Transcriptomic responses of sponge holobionts to in situ, seasonal anoxia and hypoxia. bioRxiv, 2023.02.27.530229, ver. 4 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.1101/2023.02.27.530229 Williams et al. (2008) Towards an Integrated Framework for Assessing the Vulnerability of Species to Climate Change. PLOS Biology 6(12): e325. https://doi.org/10.1371/journal.pbio.0060325 Williams et al. (2020): Research priorities for natural ecosystems in a changing global climate. Global Change Biology 26: 410–416. https://doi.org/10.1111/gcb.14856 | Transcriptomic responses of sponge holobionts to in situ, seasonal anoxia and hypoxia | Brian W Strehlow, Astrid Schuster, Warren R Francis, Lisa Eckford-Soper, Beate Kraft, Rob McAllen, Ronni Nielsen, Susanne Mandrup, Donald E Canfield | <p>Deoxygenation can be fatal for many marine animals; however, some sponge species are tolerant of hypoxia and anoxia. Indeed, two sponge species, <em>Eurypon </em>sp. 2 and <em>Hymeraphia stellifera</em>, survive seasonal anoxia for months at a ... | | Biology, Ecology, Genetics/Genomics, Invertebrates, Marine, Symbiosis | Loïc N. Michel | Maria Lopez Acosta | 2023-05-12 16:22:47 | |
19 Aug 2024
Dose, temperature and formulation shape Metarhizium anisopliae virulence against the oriental fruit fly: lessons for improving on-target control strategiesOptimizing fungal pathogen strategies for oriental fruit fly controlRecommended by Kévin Tougeron based on reviews by François Verheggen and Papa Djibril Faye
Using entomopathogenic fungi for biological control is an effective method for controlling certain crop pests, with the perspective of reducing the use of chemical pesticides. Yet, the efficiency of pathogenic fungi is dependent upon many factors that need to be evaluated to improve biological control potential in the fields (Lacey, 2001). The article by Chailleux et al. (2024) presents an exciting contribution to the field of biological pest control, specifically focusing on using entomopathogenic fungi to manage the oriental fruit fly, Bactrocera dorsalis. This fly, a member of the Tephritidae family, is a major threat to orchards in Asia, the Pacific and Africa, as it attacks fruit and causes considerable damage, in addition to having a relatively rapid biological invasion dynamic (Clarke et al. 2005). The objective of the Chailleux et al. (2024) study was to evaluate the virulence of Metarhizium anisopliae spores (strain Met69) on B. dorsalis adult flies according to various conditions: the inoculation dose and spore load, the formulation (adjuvant) and temperature conditions. The focus on host specificity and on-target applications was conducted to ensure minimal impact on non-target organisms, which is crucial for sustainable agriculture. The main challenge in this system was to achieve high strain virulence to kill wild individuals with a low number of spores—therefore limiting impact on non-target species such as natural enemies—but with a sufficient incubation period to allow transmission from mass-reared insects to wild conspecifics (Leite et al. 2022). A comparison of different inoculation methods is also provided and is interesting from a methodological point of view for future studies or even large-scale applications. Using a well-designed experimental setup, the authors show that high pathogenicity (measured by LD50) is achievable even at low spore doses and independently of the fly's sex. Lethal action speed was, however, dependent on the dose. Regarding temperature, the authors demonstrated that mycelium growth was affected by the mean temperature but, most importantly, by daily fluctuation regimes; night and day temperature alternation allowed faster growth than constant temperature. These notions of thermal fluctuations are still under-researched in terms of their modulating role in biological control yet seem central to understanding them, as the authors demonstrate here. The correlation between increased virulence and specific abiotic factors, such as temperature, offers valuable additional insights into the bioecology of the insect host and the fungal pathogen. Chailleux et al. finally point out the need for careful selection of adjuvants in formulations and pay attention to interactions with the abiotic environment to avoid compromising the effectiveness of biological control agents. Indeed, the survival rate of inoculated flies increased in the presence of the corn starch adjuvant, but this effect decreased with temperature. As corn starch unexpectedly delayed mortality, the authors suggest a potential for enhancing conspecific transmission From a broader perspective, the study emphasizes the importance of standardizing virulence evaluation to optimize biological control strategies like auto-dissemination or vectoring with sterile males, particularly in field conditions. The study contributions are timely and essential for advancing sustainable pest management strategies and improving inoculation methods. The findings underscore the need for field trials to refine these strategies, particularly in Africa, where climatic factors may affect pathogen efficacy and fly behavior. I recommend publishing this article in a referenced journal like the Peer Community Journal. References Chailleux, A. Coulibaly, ON, Diouf B, Diop S, Sohel A, Brevault T (2023) Dose, temperature and formulation shape Metarhizium anisopliae virulence against the oriental fruit fly: lessons for improving on-target control strategies. bioRxiv, ver.2 peer-reviewed and recommended by PCI Zoology https://doi.org/10.1101/2023.12.14.571642 Clarke, A. R. et al. (2005). Invasive phytophagous pests arising through a recent tropical evolutionary radiation: the Bactrocera dorsalis complex of fruit flies. Annu. Rev. Entomol., 50, 293-319. https://doi.org/10.1146/annurev.ento.50.071803.130428 Lacey, L. A. (2001). Formulation of microbial biopesticides: beneficial microorganisms, nematodes and seed treatments. J Invertebr Pathol, 77, 147. https://doi.org/10.1006/jipa.2000.5005 Leite, M. O. et al. (2022). Laboratory risk assessment of three entomopathogenic fungi used for pest control toward social bee pollinators. Microorganisms, 10, 1800. https://doi.org/10.3390/microorganisms10091800 | Dose, temperature and formulation shape Metarhizium anisopliae virulence against the oriental fruit fly: lessons for improving on-target control strategies | Anais Chailleux, Oumou N. Coulibaly, Babacar Diouf, Samba Diop, Ahmad Sohel, Thierry Brevault | <p>Entomopathogenic fungi are a promising tool for the biological control of crop pests provided low or no impact on non-target organisms. Selection for host specificity as well as on-target applications open new avenues for more sustainable stra... | | Biocontrol, Insecta, Pest management | Kévin Tougeron | 2023-12-18 11:59:30 | ||
31 Jul 2024
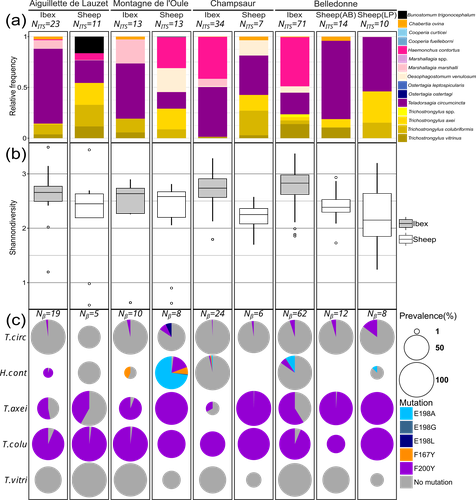
Cross-transmission of resistant gastrointestinal nematodes between wildlife and transhumant sheepWhat gets left behind? Shared nematode communities at the wildlife-livestock interface.Recommended by Karen D McCoy based on reviews by 2 anonymous reviewers
Gastrointestinal nematodes represent a major problem for livestock production across the globe, one that has intensified with the rapid and repeated evolution of multi-drug resistance (Wit et al., 2021). Understanding parasite exposure and how resistance is maintained over time are therefore of key importance for defining efficient management strategies. To date, the role wildlife play in these dynamics has been poorly studied. The work of Beaumelle et al. examine this essential question by studying the transmission dynamics of nematodes at the environmental interface between transhumant sheep and wild ungulates, more specifically with ibex (Capra ibex) that allochronically share alpine pastures when sheep are brought to graze in summer. By collecting fresh fecal material from both species and using a metabarcoding approach based on ITS-2 sequences, the authors characterise the nemabiome in each ungulate species and demonstrate that the two host species share a large portion of their parasite diversity. More importantly, by focusing on a gene (β-tubulin isotype 1) associated with resistance to a commonly used anthelmintic drug (benzimidazole), they demonstrate that both species carry resistant nematode strains, but that the diversity of strains, and particularly susceptible strains, is much higher in ibex. A key feature of the sampling design is that fecal material from both species was collected before seasonal transmission between the ungulate species could occur. Therefore, their results demonstrate that ibex are able to maintain resistant strains over long periods of time and therefore may be major nematode reservoirs for sheep infection. This important conclusion raises a series of key questions. How are resistant genotypes maintained in untreated ibex hosts? Is the cost of resistance so weak that they can coexist with susceptible strains in the absence of drug treatment or does anthelminthic contamination of the pastures maintain resistant genotypes directly in wild hosts? This work also opens several interesting perspectives: For example, what additional resistant parasites may be maintained by these wildlife hosts? What role do other wild ungulate species play in the evolution of nematode communities in transhumant sheep? An expansion of this work to the larger community of wild ungulates using alpine pastures, and an evaluation of the degree to which wild species are exposed to anthelminthic drugs released by grazing livestock into the environment is now required to understand the deeper consequences of drug treatment for shaping parasite communities and their cascading impacts for wildlife conservation, and the development of efficient and sustainable management strategies for pastoral livestock. References Beaumelle et al. Cross-transmission of resistant gastrointestinal nematodes between wildlife and transhumant sheep. bioRxiv, ver. 5 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.1101/2023.07.21.550073 Wit, J., Dilks, C.M., Andersen, E.C., 2021. Complementary Approaches with Free-living and Parasitic Nematodes to Understanding Anthelmintic Resistance. Trends Parasitol. 37, 240–250. https://doi.org/10.1016/j.pt.2020.11.008 | Cross-transmission of resistant gastrointestinal nematodes between wildlife and transhumant sheep | Camille Beaumelle, Carole Toigo, Rodolphe Papet, Slimania Benabed, Mathieu Beurier, Lea Bordes, Anais Brignone, Nadine Curt-Grand-Gaudin, Mathieu Garel, Justine Ginot, Philippe Jacquiet, Christian Miquel, Marie-Therese Poirel, Anna Serafino, Eric ... | <p>Wild and domestic ungulates can be infected with the same species of gastrointestinal parasitic nematodes. These parasites have free-living stages in the environment that contribute to the ease of transmission among different host species. In a... | | Ecology, Molecular biology, Parasitology | Karen D McCoy | 2023-07-25 10:36:28 | ||
25 Aug 2021
Up and to the light: intra- and interspecific variability of photo- and geo-tactic oviposition preferences in genus TrichogrammaNew insights into oviposition preference of 5 Trichogramma speciesRecommended by Joël Meunier based on reviews by Kévin Tougeron and Eveline C. Verhulst
Insects exhibit a great diversity of life-history traits that often vary not only between species but also between populations of the same species (Flatt and Heyland, 2011). A better understanding of the variation in these traits can be of paramount importance when it comes to species of economic and agricultural interest (Wilby and Thomas, 2002). In particular, the control of the development and expansion of agricultural pests generally requires a good understanding of the parameters that favour the reproduction of these pests and/or the reproduction of the species used to control them (Bianchi et al., 2013; Gäde and Goldsworthy, 2003). Parasitoid wasps of the genus Trichogramma are a classic example of insects involved in pest control (Smith, 1996). This genus comprises over 200 species worldwide, which have been used to control populations of a wide range of lepidopteran pests since the 1900s (Flanders, 1930; Hassan, 1993). Despite its common use, the egg-laying preference of this genus is only partially known. For example, all Trichogramma species are often thought to have positive phototaxis (or negative geotaxis) (e.g. Brower & Cline, 1984; van Atta et al., 2015), but comprehensive studies simultaneously testing this (or other) parameter among Trichogramma species and populations remain rare. This is exactly the aim of the present study (Burte et al., 2021). Using a new experimental approach based on automatic image analysis, the authors compared the photo- and geo-tactic oviposition preference among 5 Trichogramma species from 25 populations. Their results first confirm that most Trichogramma species and populations prefer light to shade, and higher to lower positions for oviposition. Interestingly, they also reveal that the levels of preference for light and gravity show inter- and intraspecific variation (probably due to local adaptation to different strata) and that both preferences tend to relax over time. Overall, this study provides important information for improving the use of Trichogramma species as biological agents. For example, it may help to establish breeding lines adapted to the microhabitat and/or growing parts of plants on which agricultural pests lay eggs most. Similarly, it suggests that the use of multiple strains with different microhabitat selection preferences could lead to better coverage of host plants, as well as a reduction in intraspecific competition in the preferred parts. Finally, this study provides a new methodology to efficiently and automatically study oviposition preferences in Trichogramma, which could be used in other insects with a particularly small size. References Bianchi, F. J. J. A., Schellhorn, N. A. and Cunningham, S. A. (2013). Habitat functionality for the ecosystem service of pest control: reproduction and feeding sites of pests and natural enemies. Agricultural and Forest Entomology, 15, 12–23. https://doi.org/10.1111/j.1461-9563.2012.00586.x Burte V., Perez G., Ayed F., Groussier G., Mailleret L, van Oudenhove L. and Calcagno V. (2021). Up and to the light: intra- and interspecific variability of photo-and geo-tactic oviposition preferences in genus Trichogramma. bioRxiv, 2021.03.30.437671, ver. 4 peer-reviewed and recommended by PCI Zoology. https://doi.org/10.1101/2021.03.30.437671 Brower, J. H. and Cline, L. D. (1984). Response of Trichogramma pretiosum and T. evanescens to Whitelight, Blacklight or NoLight Suction Traps. The Florida Entomologist, 67, 262–268. https://doi.org/10.2307/3493947 Flanders, S. E. (1930). Mass production of egg parasites of the genus Trichogramma. Hilgardia, 4, 465–501. https://doi.org/10.3733/hilg.v04n16p465 Flatt, T. and Heyland, A. (2011). Mechanisms of life history evolution: the genetics and physiology of life history traits and trade-offs. Oxford University Press. https://doi.org/10.1093/acprof:oso/9780199568765.001.0001 Gäde, G. and Goldsworthy, G. J. (2003). Insect peptide hormones: a selective review of their physiology and potential application for pest control. Pest Management Science, 59, 1063–1075. https://doi.org/10.1002/ps.755 Hassan, S. A. (1993). The mass rearing and utilization of Trichogramma to control lepidopterous pests: Achievements and outlook. Pesticide Science, 37, 387–391. https://doi.org/10.1002/ps.2780370412 Smith, S. M. (1996). Biological Control with Trichogramma : Advances, Successes, and Potential of Their Use. Annual Review of Entomology, 41, 375–406. https://doi.org/10.1146/annurev.en.41.010196.002111 van Atta, K. J., Potter, K. A. and Woods, H. A. (2015). Effects of UV-B on Environmental Preference and Egg Parasitization by Trichogramma Wasps (Hymenoptera: Trichogrammatidae). Journal of Entomological Science, 50, 318–325. https://doi.org/10.18474/JES15-09.1 Wilby, A. and Thomas, M. B. (2002). Natural enemy diversity and pest control: patterns of pest emergence with agricultural intensification. Ecology Letters, 5, 353–360. https://doi.org/10.1046/j.1461-0248.2002.00331.x | Up and to the light: intra- and interspecific variability of photo- and geo-tactic oviposition preferences in genus Trichogramma | Burte, V., Perez, G., Ayed, F. , Groussier, G., Mailleret, L., van Oudenhove, L. and Calcagno, V. | <p>Trichogramma are parasitic microwasps much used as biological control agents. The genus is known to harbor tremendous diversity, at both inter- and intra-specific levels. The successful selection of Trichogramma strains for biocontrol depends o... | | Behavior, Biocontrol, Biodiversity, Ecology, Insecta, Parasitology, Pest management, Systematics, Terrestrial | Joël Meunier | Kévin Tougeron, Eveline C. Verhulst | 2021-04-02 16:10:28 | |
21 Jun 2023
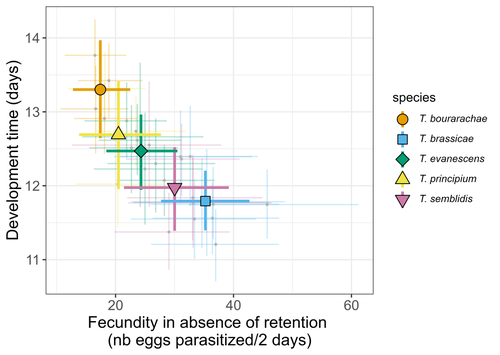
Life-history traits, pace of life and dispersal among and within five species of Trichogramma wasps: a comparative analysisThe relationship between dispersal and pace-of-life at different scalesRecommended by Jacques Deere based on reviews by Mélanie Thierry and 1 anonymous reviewerThe sorting of organisms along a fast-slow continuum through correlations between life history traits is a long-standing framework (Stearns 1983) and corresponds to the pace-of-life axis. This axis represents the variation in a continuum of life-history strategies, from fast-reproducing short-lived species to slow-reproducing long-lived species. The pace-of-life axis has been the focus of much research largely in mammals, birds, reptiles and plants but less so in invertebrates (Salguero-Gómez et al. 2016; Araya-Ajoy et al. 2018; Healy et al. 2019; Bakewell et al. 2020). Outcomes from this research have highlighted variation across taxa on this axis and mixed support for, and against, patterns expected of the pace-of-life continuum. Given this, a greater understanding of the variation of the pace-of-life across-, and within, taxa are needed. Indeed, Guicharnard et al. (2023) highlight several points regarding our broader understanding of pace-of-life. In general, invertebrates are poorly represented, the variation of pace-of-life across taxonomic scales is less well understood and the relationship between pace-of-life and dispersal, a key life history, requires more attention. Here, Guicharnard et al. (2023) provide a first attempt at addressing the relationship between dispersal and pace-of-life at different scales. The authors, under controlled conditions, investigated how life-history traits and effective dispersal covary for 28 lines from five species of endoparasitoid wasps from the genus Trichogramma. At the species level negative correlations were found between development time and fecundity, matching pace-of-life axis predictions. Although this correlation was not found to be significant among lines, within species, a similar pattern of a negative correlation was observed. This outcome matches previous findings that consistent pace-of-life axes become more difficult to find at lower taxonomic levels. Unlike the other life-history traits measured, effective dispersal showed no evidence of differences between species or between lines. The authors also found no correlation between effective dispersal and other-life history traits which suggests no dispersal/life-history syndromes in the species investigated. One aspect that was not assessed was the impact of density dependence on pace-of-life and effective dispersal, largely as this was a first step in assessing relationship of dispersal with pace-of-life at different scales. However, the authors do acknowledge the importance of future studies incorporating density dependence and that such studies could potentially lead to more generalizable understanding of pace-of-life and dispersal within Trichogramma. A pleasant addition was the link to potential implications for biocontrol. This addition showed an awareness by the authors of how insights into pace-of-life can have an applied component. The results of the study highlighted that selecting for specific lines of a species, to maximise a trait of interest at the cost of another, may not be as effective as selecting different species when implementing biocontrol. This is especially important as often single, established species used in biocontrol are favoured without consideration of the potential of other species which can lead to more efficient biocontrol. REFERENCES Araya-Ajoy, Y.G., Bolstad, G.H., Brommer, J., Careau, V., Dingemanse, N.J. & Wright, J. (2018). Demographic measures of an individual's "pace of life": fecundity rate, lifespan, generation time, or a composite variable? Behavioral Ecology and Sociobiology, 72, 75. | Life-history traits, pace of life and dispersal among and within five species of *Trichogramma* wasps: a comparative analysis | Chloé Guicharnaud, Géraldine Groussier, Erwan Beranger, Laurent Lamy, Elodie Vercken, Maxime Dahirel | <p>Major traits defining the life history of organisms are often not independent from each other, with most of their variation aligning along key axes such as the pace-of-life axis. We can define a pace-of-life axis structuring reproduction and de... | | Biology, Ecology, Insecta, Invertebrates, Life histories | Jacques Deere | 2023-01-25 18:15:20 | ||
09 Feb 2023

A novel nematode species from the Siberian permafrost shares adaptive mechanisms for cryptobiotic survival with C. elegans dauer larvaA novel nematode species from the Siberian permafrost shares adaptive mechanisms for cryptobiotic survival with C. elegans dauer larvaRecommended by Isa Schon based on reviews by 3 anonymous reviewersThis article [1] investigated two nematode genera, Panagrolaimus and Plectus, from the Siberian permafrost to unravel the adaptations allowing them to survive cryptobiosis; radio carbon dating showed that the individuals of Panagrolaimus had been in cryobiosis in Siberia for as long as 46,000 years! I was impressed by the multidisciplinary approach of this study, including morphological as well as phylogenetic and -genomic analyses to describe a new species. In triploids as some of the species studied here, it is quite challenging to assemble a novel genome. The authors furthermore not only managed to successfully reanimate the Siberian specimens but could also expose them to repeated freezing and desiccation in the lab, not an easy task. This study reports some amazing discoveries - comparing the molecular toolkits between C. elegans and Panagrolaimus and Plectus revealed that several components were orthologues. Likewise, some of the biochemical mechanisms for surviving freezing in the lab turned out to be similar for C. elegans and the Siberian nematodes. This study thus provides strong evidence that nematodes developed specific mechanisms allowing them to stay in cryobiosis over very long times. A surprising additional experimental result concerns the well-studied C. elegans - dauer larvae of this species can stay viable much longer after periods of animated suspension than previously thought. I highly recommend this article as it is an important contribution to the fields of evolution and molecular biology. This study greatly advanced our understanding of how nematodes could have adapted to cryobiosis. The applied techniques could also be useful for studying similar research questions in other organisms. Reference [1] Shatilovich A, Gade VR, Pippel M, Hoffmeyer TT, Tchesunov AV, Stevens L, Winkler S, Hughes GM, Traikov S, Hiller M, Rivkina E, Schiffer PH, Myers EW, Kurzchalia TV (2023) A novel nematode species from the Siberian permafrost shares adaptive mechanisms for cryptobiotic survival with C. elegans dauer larva. bioRxiv, 2022.01.28.478251, ver. 6 peer-reviewed and recommended by Peer Community in Zoology. https://doi.org/10.1101/2022.01.28.478251 | A novel nematode species from the Siberian permafrost shares adaptive mechanisms for cryptobiotic survival with C. elegans dauer larva | Anastasia Shatilovich, Vamshidhar R. Gade, Martin Pippel, Tarja T. Hoffmeyer, Alexei V. Tchesunov, Lewis Stevens, Sylke Winkler, Graham M. Hughes, Sofia Traikov, Michael Hiller, Elizaveta Rivkina, Philipp H. Schiffer, Eugene W Myers, Teymuras V. K... | <p style="text-align: justify;">Some organisms in nature have developed the ability to enter a state of suspended metabolism called cryptobiosis1 when environmental conditions are unfavorable. This state-transition requires the execution of comple... | | Ecology, Evolution, Genetics/Genomics | Isa Schon | 2022-05-20 14:32:02 |




